
Arxius de Miscel·lània Zoològica. Volumen 17 (2019) Páginas: 73-84
Biocontrol of the pine processionary moth Thaumetopoea pityocampa (Denis and Schiffermuller, 1775) with plant extracts
Kherroubi, M., Mouhouche, F., Zerrouk Izzeddine, Z., Chahbar, M.
DOI: https://doi.org/10.32800/amz.2019.17.0073Descargar
PDFCita
Kherroubi, M., Mouhouche, F., Zerrouk Izzeddine, Z., Chahbar, M., 2019. Biocontrol of the pine processionary moth Thaumetopoea pityocampa (Denis and Schiffermuller, 1775) with plant extracts. Arxius de Miscel·lània Zoològica, 17: 73-84, DOI: https://doi.org/10.32800/amz.2019.17.0073-
Fecha de recepción:
- 15/03/2019
-
Fecha de aceptación:
- 06/06/2019
-
Compartir
-


-
Visitas
- 4182
-
Descargas
- 1096
Abstract
Biocontrol of the pine processionary moth Thaumetopoea pityocampa (Denis and Schiffermuller, 1775) with plant extracts
The purpose of this study was to propose alternative solutions based on the use of natural, bio-insecticidal products to fight against the processionary moth, Thaumetopoea pityocampa (T. pityocampa), considered a defoliator because of the damage it causes to silvicultural. This insect also causes health problems in forests or wooded areas open to the public. To meet this objective, we evaluated the toxicity of six extracts from two plants: Taxus baccata and Populus nigra, collected from the regions of Blida, against first instar larvae of T. pityocampa. The results from these biological tests showed a significant difference between the six extracts. The LD50 (lethal dose) values were calculated by probit analysis using Finney’s table. The LD50 was also low for the two extracts Populus Methanol and Populus Methanol Chloroform (1.4 mg/ml and 3.3 mg/ml, respectively). However, the LD50 was relatively high for the rest of the extracts. In conclusion, the Bio-insecticides of Populus Methanol (PMP) and Populus Methanol Chloroform (PMC) showed to be active and could be integrated into the biological control of the egg parasitoids of T. pityocampa.
Key words: Thaumetopoea pityocampa, LD50, Populus nigra, Taxus baccata
Resumen
Control biológico de la mariposa de la procesionaria del pino Thaumetopoea pityocampa (Denis y Schiffermuller, 1775) mediante varios extractos vegetales
La finalidad del presente estudio es ofrecer alternativas con bioinsecticidas elaborados a partir de productos naturales para combatir la mariposa de la procesionaria, Thaumetopoea pityocampa (T. pityocampa), considerada una plaga defoliadora debido a los daños que causa en la silvicultura. Este insecto ocasiona asimismo problemas de salud en bosques o zonas forestales abiertas al público. Para ello hemos evaluado la toxicidad de seis extractos procedentes de dos plantas: Taxus baccata y Populus nigra, recogidos en distintas zonas de Blida, contra larvas en estadio 1 de T. pityocampa. Los ensayos biológicos han demostrado una reducción significativamente diferente de estos insectos con cada uno de los seis extractos. Los valores LD50 (dosis letal) se calcularon aplicando el análisis probit con la tabla de Finney. Los valores LD50 fueron bajos en el caso de dos extractos: el metanol de chopo (Populus Methanol) y el cloroformo-metanol de chopo (Populus Methanol Chloroform), con 1,4 mg/ml y 3,3 mg/ml, respectivamente. En cambio, las dosis LD50 del resto de extractos fueron relativamente elevadas. En conclusión, los insecticidas biológicos metanol de chopo (PMP) y cloroformo-metanol de chopo (PMC) resultaron más eficaces y, por lo tanto, podrían integrarse en el control biológico de T. pityocampa junto con parasitoides de huevos.
Palabras clave: Thaumetopoea pityocampa, LD50, Populus nigra, Taxus baccata
Resum
Control biològic de la papallona de la processionària del pi Thaumetopoea pityocampa (Denis i Schiffermuller, 1775) mitjançant diversos extractes vegetals
La finalitat del present estudi és oferir alternatives amb bioinsecticides elaborats a partir de productes naturals per combatre la papallona de la processionària, Thaumetopoea pityocampa (T. pityocampa), considerada una plaga defoliadora a causa dels danys que produeix en la silvicultura. Aquest insecte ocasiona així mateix problemes de salut en boscos o zones forestals obertes al públic. Amb aquest objectiu hem avaluat la toxicitat de sis extractes procedents de dues plantes: Taxus baccata i Populus nigra, recollits en diferents zones de Blida, contra larves en estadi 1 de T. pityocampa. Els assajos biològics han demostrat una reducció significativament diferent d’aquests insectes amb cadascun dels sis extractes. Els valors LD50 (dosi letal) es van calcular aplicant l’anàlisi probit amb la taula de Finney. Els valors LD50 van ser baixos en el cas de dos extractes: el metanol de pollancre (Populus Methanol) i el cloroform-metanol de pollancre (Populus Methanol Chloroform), amb 1,4 mg/ml i 3,3 mg/ml, respectivament. En canvi, les dosis LD50 de la resta d’extractes van ser relativament elevades. En conclusió, els insecticides biològics metanol de pollancre (PMP) i cloroform-metanol de pollancre (PMC) van resultar més eficaços i, per tant, es podrien integrar en el control biològic de T. pityocampa juntament amb parasitoides d’ous.
Paraules clau: Thaumetopoea pityocampa, LD50, Populus nigra, Taxus baccata
Introduction
Forest areas are estimated to cover about four billion hectares worldwide, including 30 % of the land area (FAO, 2015). In Algeria, forest covers an area of 4.7 million hectares with an afforestation rate of 11 % (DGF, 2007). Cedar forest is part of the Mediterranean ecosystems and has a fundamental role in the equilibrium of the mountain ecosystem in North Africa. In Algeria, the Atlas cedar, Cedrus atlantica Manetti (1844), occupies 40,000 ha. Its great biological richness is a focus of exceptional interest in regards to protection and extension. The requirement to protect this cedar is always upstream of forestry programs. These biotic communities are threatened by various species of defoliators, mostly the pine processionary moth, Thaumetopoea pityocampa (Fraval, 2007). This primary insect weakens its host and exposes it to other insects, in particular, xylophages (Zamoum, 2002).
The larvae of Thaumetopoea pityocampa, known for their gregarious lifestyles, their voracity, their mode of moving in a single file (Seigue, 1985) and their urtication properties, cause many health problems for humans and animals (Battisti et al., 2017). The extension of the range of the host species plays an important role in the movement of this insect (Leblond, 2009). This situation of progradation necessitates local control actions, especially in forests or wooded areas used by the public, in order to reduce infestations of this pest and to protect the most sensitive stands, particularly: (a) sensitive young stands in protected forest and production forest when the survival of trees is threatened by strong repeated defoliation; and (b) forests frequented by the public, where the control decisions differs as the primary damage is the unsightly trees in the leafless period and the harmful reactions due to urtication caused by the hair of the larvae.
Plants produce secondary compounds to defend themselves against organisms such as pathogens and pests (Meric, 2005). These secondary compounds have a wide mode of action on insects, including repulsive effects and digestive inhibition (Regnault-Roger and Hamraoui, 1997). Historically, botanicals were used long before other kinds of pesticides. The ancient Chinese, Greeks, and Romans used plants or plant extracts with sulphur and arsenic as insecticides (Tschirley, 1979). More than 2,000 plant species with insecticidal properties have been recorded (Grainge and Ahmed, 1988). It has been reported that the Romans used powders prepared from Veratrum sp. as insecticides and rodenticides. In the tropics the use of neem (Azadirachta indica Juss. Meliaceae) has been recorded for at least 4,000 years (Larson, 1989). In the 19th century, only a few compounds of plant origin were identified and widely used as repellents or toxic products, including nicotine (alkaloid) and its derivatives, rotenone, pyrethrum, and vegetable oils. Several studies have shown the importance of Taxus baccata and Populus nigra. Among these studies, we can mention those of Roberts and Wink (1998) on two new European yew taxoids (Taxus baccata) that act as pyrethroid insecticides Synergists against the black vine weevil (Otiorhynchus sulcatus). The seeds and foliage of Taxus baccata are rich in toxic alkaloids (Miller, 1980). All parts of the tree are toxic and contain Taxine, a complex mixture of alkaloids that are rapidly absorbed by digestion and interfere with the action of the heart (Cooper and Johnson, 1984). In addition, Popova et al. (2005) evaluated the antibacterial activity of black propolis from Turkey and its qualitative and quantitative chemical composition. The effects insecticides of Populus nigra has been evaluated by Cao et al. (2010) against Lymantria dispar L.
The objective of our work was to study the effects of the application of plant extracts against the processionary moth, T. pityocampa. We assessed the insecticide potential and disturbance of the host recognition by natural extracts. This effect was measured on several biological parameters that characterize the dynamics of the insect population in general and the gradation in particular while preserving the parasitoids.
Material and methods
Harvesting and preparation of the biological material
During the summer of 2018, we harvested a 200 egg mass of the processionary moth, T. pityocyampa. The eggs were stored individually in test tubes of 10 cm in length and 1 cm in diameter. The 1st instar larvae (L1) of T. pityocampa were collected from infected Cedrus atlantica trees in the forest areas (Chréa) in the Blida region of Algeria at an altitude of 1,388 m.
Plant material
The aerial parts (stem and leaves) of Taxus baccata and Populus nigra were collected during April 2018 at a floral stage of the plant, in the region of Blida, 50 km southwest of Algiers (Algeria) (36° 26′ 4.15″ N 2° 53′ 20.29″ E) (fig. 1). Plant samples were dried naturally on laboratory benches at room temperature (24–25 ºC) for two months. The dried plant materials were ground to a homogenous powder using a RECOCHET blender (Réf. CSL04).
Fig. 1. Location map of Blida (study station) Fig. 1. Mapa de localización de Blida (estación de estudio)
Preparation of plant total extracts
To prepare the total extracts from the two plants species, 10 g of each powder was shaken separately for 48 h in a 100 ml volume (V) of: Pure Methanol, Water-Methanol (Volume1/Volume2) and Methanol-Chloroform (2V1/V2) at room temperature (25 ºC). The extracts were filtered using Whatman filter paper (No. 1). Each filtrate was concentrated under reduced pressure at 40ºC using a Rotavapor (Buchi R-215). Each extract was resuspended in methanol to make a stock solution. The extracts were kept at 4 ºC until further tests. Finally, six extracts were prepared: Taxus Methanol Pure (TMP), Taxus Methanol Chloroform (TMC), Taxus Methanol-Water (TME), Populus Methanol Pure (PMP), Populus Methanol Chloroform (PMC),and Populus Methanol-Water (PME).
Rate of extraction yield in percentages
The performance of the extraction of each fraction (R) is defined as a ratio of extract quantity (E) on the quantity of plant material (MS):
R = E/MS x 100 (Abbott, 1925)
Insecticidal effect of plant extracts
The insecticidal effect of the extracts against the first instar larvae of T. pityocampa was evaluated by means of contact toxicity. For this purpose, we used an impregnated paper assay, following the method described by Stefanazzi et al. (2011) with some modifications. Based on preliminary results, a range of doses: 0.2 % (2 mg/ml), 0.4 % (4 mg/ml) and 0.8 % (8 mg/ml) were formulated and applied to Whatman filter paper (No. 1) (9 cm diameter). The extracted solvent was air-dried for 10 min. Each filter paper was then placed inside a glass Petri dish with 20 larvae of this insect and 15 g amounts of fresh needles of Pinus pinea. The dishes were kept in optimal laboratory condition (27 ± 2 ºC, 70 ± 5 % relative humidity and in darkness). Following exposure, the mortality larvae were determined at 24 h, 48 h and 72 h. Petri dishes with sterile water (T0) were used as controls. Three replicates were used for each dose and exposure time combination, and larvicidal activity of the plant extracts was expressed as the percentage of mean mortality of the larvae. The lethal dose (LD50) values were calculated by probit analysis using Finney’s table.
Statistical analyses
Statistical analysis was conducted using SAS software 9. The differences between treatments were detected by analysis of variance (ANOVA). P-values < 0-05 were regarded as significant. The multiple comparisons (two-two) of the averages between the different plant extracts were performed using Tukey’s test of least significant differences (LSD).
Results and discussion
Rate (%) of yield extraction
Table 1 shows the yields of the crude extracts obtained from aerial parts of two local plants, Taxus baccata and Populus nigra, using different solvents (Methanol, Methanol-Chloroform and Methanol-Water). The extraction yield was in the range of 20 %-77 %, with the highest yield being obtained for Populus nigra when the solvents Methanol and Methanol-Chloroform gave the best extraction results (61 % and 77 % respectively).

Table 1. Yields of extract solvents from Taxus baccata and Populus nigra. Tabla 1. Rendimiento de los extractos solventes obtenidos a partir de Taxus baccata y Populus nigra.
Descriptive analysis of mortality average
Forty-eight hours after the application of the compounds tested at different concentrations against the 1st instar larvae of T. pityocampa (fig. 2, table 2), we found that the extracts PME and PMC were highly toxic, causing mortality averages, respectively, of 50 % and 35 % for dose 1 (D1) (2 mg/ml), 88.35% and 40 % for dose 2 (D2) (4 mg/ml), and 100 % and 95 % for dose 3 (D3) (8 mg/ml). In contrast, extracts PME, TMP, TMC and TME had low toxicity against the 1st instar larvae of T. pityocampa. Based on these results, D3 showed important efficiency after 48 h in accordance with D1 and D2 and with a mortality average of 100 % and 95 %, respectively, for the PMP extract and PMC extract. Dose D3 caused 50 % of mortality for the extracts, PME, TMC, TMP and TME. Furthermore, D1 and D2 did not reach 50 % of mortality until 48 h of exposure time. The extract PMP reached 70 % and 88.35 % respectively for D1 and D2. In the same context, Kesdek et al. (2014) evaluated the larvicidal activity of some plant extracts against the second larvae of the processionary moth, T. pityocampa in Turkey. They found a minimal mortality average of 20 % after 24 h of exposure to the extracts of Origanum onites and Satureja hortensi, contrary to the maximum rate of 73.33 % for the extracts of Tanacetum argyrophyllum and Nepeta meyeri.
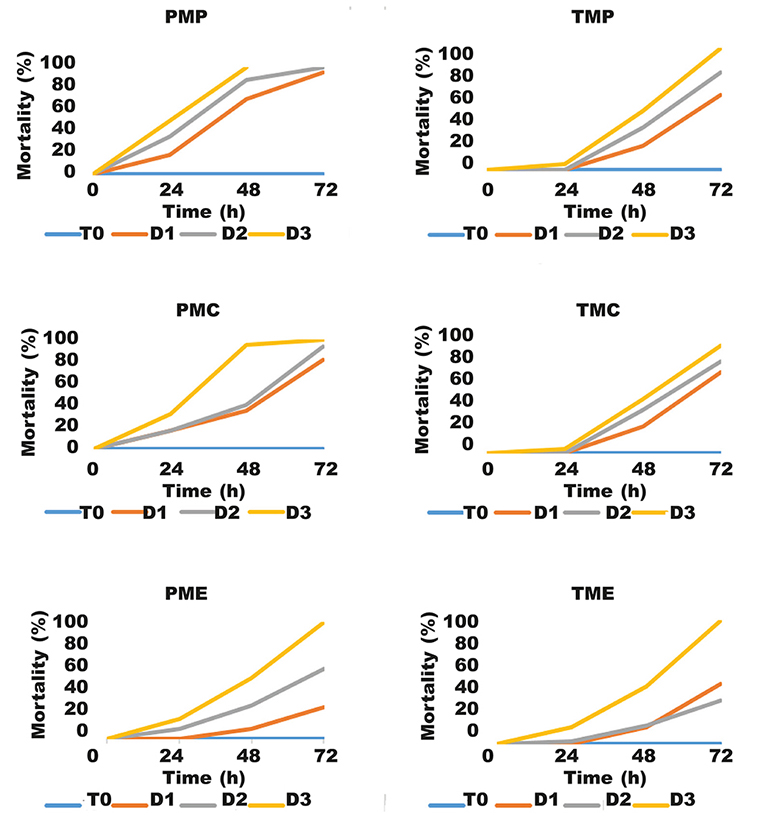
Fig. 2. Evaluation of 1st instar larvae of Thaumetopoea pityocampa after treatment with the extracts TMP, TMC, TME, PMP, PME, PMC: T0, witness of our experimentation (by sterile water); D, extract dose. Fig. 2. Evaluación de larvas en estadio 1 de Thaumetopoea pityocampa tras el tratamiento con extractos de TMP, TMC, TME, PMP, PME, PMC: T0, testigo de nuestro experimento (con agua estéril); D, dosis de extracto.

Table 2. Descriptive analysis of the mortality averages (48 h): TMP, Taxus Methanol Pure; TMC, Taxus Methanol Chloroform; TME, Taxus Methanol–Water; PMP, Populus Methanol Pure; PMC, Populus Methanol Chloroform; PME, Populus Methanol–Water. Tabla 2. Análisis descriptivo de la medias de mortalidad (48 h): TMP, metanol de tejo puro; TMC, cloroformo–metanol de tejo; TME, metanol de tejo–agua; PMP, metanol de chopo puro; PMC, cloroformo–metanol de chopo; PME, metanol de chopo–agua.
Variation of the mortality rate
Table 3 summarizes the daily cumulative mortality rate of L1 after the exposure time of 1st instar larvae (L1) of T. pityocampa on different concentrations of the plant extracts from Taxus baccata and Populus nigra. The ANOVA procedure shows a highly significant difference between the variables, days (DF = 2, F = 350, P < 0.0001), extracts (DF = 4, F = 33.10, P < 0.0001) and dose (DF = 2, F = 67.17, P < 0.0001) that are in accordance with the mortality rate.
Table 3. Variance analyses of Thaumetopoea pityocampa mortality. The F–value is the ratio produced by dividing the Mean Square for the Model by the Mean. The Type III SS is the sum of squares for a balanced test of each effect, adjusted for all other effects. (DF is the degrees of freedom.) Tabla 3. Análisis de varianza de la mortalidad de Thaumetopoea pityocampa. El valor F es la relación existente tras dividir la media cuadrática del modelo por la media. El tipo III SS es la suma de los cuadrados para un test de equilibrado de cada efecto, ajustada para cada uno de los otros efectos. (DF son los grados de libertad.)
Comparison of the averages
The multiple comparisons (two-two) of the averages between the different plant extracts using the test of least significant differences (LSD) show no significant difference between extracts TMC and TMP, TMP and PME, TME and PME. In contrast, we found a significant difference of 0.05 between the remaining comparisons.
The grouping of homogeneous extracts revealed five groups, ranging from highly effective to less effective depending on the average mortality of larvae L1 of T. pityocampa (table 4). Extract PMP was found in group A. This extract has greater toxicity than all the other extracts. Group B represented the extract PMC. Groups C and D respectively represented the extracts PME and TME. Finally, an intermediate group, CD, consisted of both TMC and TMP extracts. These results confirm the study of Yang et al. (2003) about the importance of poplars against the defoliating insects. Their bio-assays of transgenic poplar clone 741 on the larvae of Lymantria dispar, Clostera anachoreta (Fabricius) were carried out annually. Their results showed that three transgenic sub-clones tested had high resistance against these insects. The insecticidal activity on the first larvae stage was obvious and decreased gradually with the development of the larvae (instar stage). Growth and development of the surviving larvae were seriously inhibited and delayed and in some cases could not even complete their development.

Table 4. Multiple comparisons (two-two) of plants extracts by Tukey test (LSD): Nb, number of observations; TMP, Taxus Methanol Pure; TMC, Taxus Methanol Chloroform; TME, Taxus Methanol–Water; PMP, Populus Methanol Pure; PMC, Populus Methanol Chloroform; PME, Populus Methanol–Water. Tabla 4. Comparaciones múltiples (dos-dos) de los extractos vegetales mediante la prueba de Tukey (LSD): Nb, número de observación; TMP, metanol de tejo puro; TMC, cloroformo–metanol de tejo; TME, metanol de tejo–agua; PMP, metanol de chopo puro; PMC, cloroformo–metanol de chopo; PME, metanol de chopo–agua.
LD50 of the various substances tested against the 1st instar larvae L1 of Thaumetopoea pityocampa
Table 5 and figure 3 summarize the lethal amounts with 50% (LD50) of the extracts of Taxus baccata and Populus nigra.

Table 5. LD50 values of the different extracts of Taxus baccata and Populus nigra against 1st instar larvae (L1) of Thaumetopoea pityocampa: LD50, lethal dose causes the death of 50 % of a group of test insect; R2, Coefficient of determination; TMP, Taxus Methanol Pure; TMC, Taxus Methanol Chloroform; TME, Taxus Methanol–Water; PMP, Populus Methanol Pure; PMC, Populus Methanol Chloroform; PME, Populus Methanol–Water. Tabla 5. Valores LD50 de los diferentes extractos de Taxus baccata y Populus nigra para larvas en estado 1 (L1) de Thaumetopoea pityocampa: LD50, la dosis letal causa la muerte del 50 % de un grupo de insectos sometido a ensayo; R2, coeficiente de determinación; TMP, metanol de tejo puro; TMC, cloroformo–metanol de tejo; TME, metanol de tejo–agua; PMP, metanol de chopo puro; PMC, cloroformo–metanol de chopo; PME, metanol de chopo–agua.
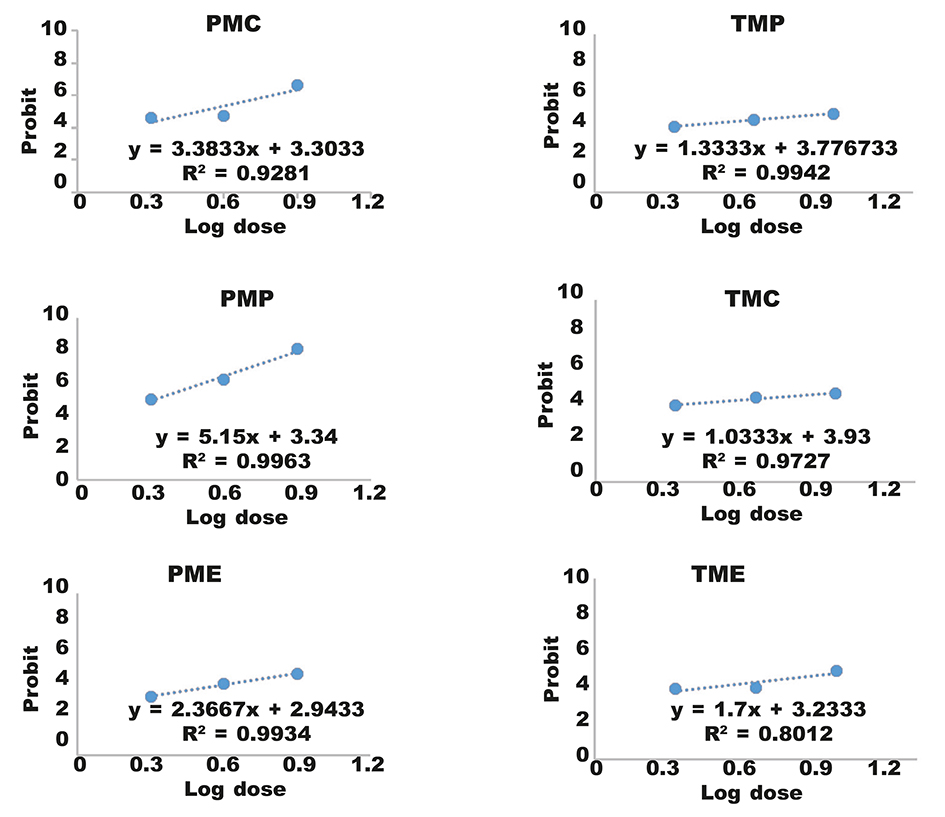
Fig. 3. Right regression of the relation log dose–probit analysis. Fig. 3. Regresión adecuada de la relación dosis logarítmica–probit.
LD50 results for the 1st instar larvae (L1) of T. pityocampa showed that two of the plant extracts tested were interesting in terms of toxicity: the PMP extract and the PMC extract. They presented the weakest concentration of LD50 at 0.18 % (1.8 mg/ml) and 0.37 % (3.7 mg/ml) respectively. On the other hand, extracts PME, TMP, TMC and TME were less toxic, with values respectively of 0.75 % (7.5 mg/ml), 0.8 % (8 mg/ml), 0.93% (9,3 mg/ml) and 0.88 % (8.8 mg/ml). Kesdek et al. (2014) found a mortality rate between 3.33 % and 86.6 % for Origanum rotundifolium extracts and Nepeta meyeri extracts against the 2nd, 3rd and 4th instar larvae of T. pityocampa. In addition, Ouakid et al. (2005) studied the effects of some plant extracts that associated Oak cork against Lymantria dispar; the results showed that the extracts of Quercus suber and Quercus faginea were most effective on the old larvae of Lymantria dispar. Charrière-Ladreix (1973) studied the flavonoid secretion of Populus nigra L. buds. The same author in 1976 showed the intracellular distribution of the flavonic secretion of Populus nigra L. Salle and Andary (1990) presented the importance of polyphenols on the resistance mechanisms of four poplar cultivars in response to Viscum album L. attack. In addition, the anti-xanthine oxidase and anti-radical activity of Populus nigra bud extracts were evaluated by Adrar (2009). Furthermore, studies by Daniewski et al. (1998) highlight the insecticidal importance and the high anti-nutritional activity of extracts of Taxus baccata against the pests Tribolium confusum, Trogoderma granarium and Sitophilus granarius. Elsohly et al. (1997) reported the taxane content of needle stems of Taxus baccata from nurseries in the United States.
Conclusion
Findings from our study indicate that extracts of Taxus baccata and Populus nigra had a larvicidal effect on T. pityocampa. The extracts Populus Methanol Pure and Populus Methanol Chloroform show promising insecticidal activities. These plant extracts can therefore be considered potential alternatives to control against the instar larvae of T. pityocampa. Further research should be conducted to isolate the bio-compounds involved in this activity and to determine their mode of action in order to facilitate the development of formulations of new bio-pesticides in the control of T. pityocampa. Planning and performing tests under natural forest conditions is needed to evaluate the real effectiveness of these extracts against T. pityocampa.
Acknowledgements
This study was funded by the Ministry of High Education and Scientific Research, the National High School of Agronomy, Ecole Normale Superieure Kouba, and the PhD program, Algeria. Thanks go to the head and laboratory staff at of the Zoology Department, ENSA Algiers and eco-biology laboratory-ENS-Kouba Algeria for providing laboratory facilities.
















