
Arxius de Miscel·lània Zoològica. Volumen 17 (2019) Páginas: 59-71
Population fluctuations of Gynaikothrips ficorum (Marchal, 1908) (Thysanoptera, Tubulifera) and natural enemies on leaf gall of Ficus retusa in Algeria
Ziouani, K., Benzahra, A., Saharaoui, L.
DOI: https://doi.org/10.32800/amz.2019.17.0059Descargar
PDFCita
Ziouani, K., Benzahra, A., Saharaoui, L., 2019. Population fluctuations of Gynaikothrips ficorum (Marchal, 1908) (Thysanoptera, Tubulifera) and natural enemies on leaf gall of Ficus retusa in Algeria. Arxius de Miscel·lània Zoològica, 17: 59-71, DOI: https://doi.org/10.32800/amz.2019.17.0059-
Fecha de recepción:
- 11/03/2019
-
Fecha de aceptación:
- 04/06/2019
-
Fecha de publicación:
- 30/06/2019
-
Compartir
-


-
Visitas
- 4827
-
Descargas
- 988
Abstract
Population fluctuations of Gynaikothrips ficorum (Marchal, 1908) (Thysanoptera, Tubulifera) and natural enemies on leaf gall of Ficus retusa in Algeria
Monthly fluctuations of thrip populations on Ficus retusa were monitored and parasites and predators in Boudouaou, Algeria, were identified. Twenty leaves were collected fortnightly from 30 trees from January to December 2015. The number of eggs increased significantly in August with 11,163 eggs and was slightly higher in October with 11,471 eggs. Presence was rare in spring and winter. The 1st and 2nd larval stages (L1 and L2) overlapped: L1 were present from August to October with 27 and 51individuals respectively, and L2 were present from August to December with 65 to 186 individuals respectively. Prepupae were active in April and June, reaching 46 and 50 individuals respectively, before increasing to 266 in November. Highest numbers of pupae were observed in June (171) and July (135). The number of adults gradually increased, reaching a maximum of 642 in early December. Two predators were identified: Montandoniola confusa (Hemiptera, Anthocoridae) and Nephus peyerimhoffi (Coleoptera, Coccinellidae). A parasite, Tetrastichus gentilei (Hymenoptera, Eulophidae), was also later identified.
Key words: Gynaikothrips ficorum, Ficus retusa, Predators, Parasite, Algeria
Resumen
Fluctuaciones en las poblaciones de Gynaikothrips ficorum (Marchal, 1908) (Thysanoptera, Tubulifera) y de sus enemigos naturales en las agallas de hojas de Ficus retusa en Argelia
El estudio consiste en la observación de las fluctuaciones mensuales de las poblaciones de arañuelas en Ficus retusa, así como de sus parásitos y depredadores conocidos, en Boudouaou. Para ello, cada quince días se recogieron 20 hojas de cada uno de los árboles seleccionados, durante 12 meses de enero a diciembre de 2015. Aparentemente, el número de huevos aumenta considerablemente en agosto, cuando se registraron 11.163 huevos, valor que se superó ligeramente en octubre, con 11.471 huevos, frente a una ausencia casi total de los mismos en primavera e invierno. Se solapan los estadios larvarios 1 y 2 (L1 y L2): L1 va de agosto a octubre, con 27 y 51 individuos respectivamente, seguido de L2, con 65 y 186 individuos de octubre a diciembre. Las prepupas están activas en abril y junio, cuando alcanzan 46 y 50 individuos, respectivamente, para aumentar hasta 266 en noviembre, si bien muestran valores elevados en junio y julio, con 171 y 135 ejemplares respectivamente. Los individuos adultos aumentan progresivamente hasta llegar al máximo a principios de diciembre, con 642 individuos. Se han identificado asimismo, dos depredadores: Montandoniola confusa (Hemiptera, Anthocoridae) y Nephus peyerimhoffi (Coleoptera, Coccinellidae) y solo un parásito: Tetrastichus gentile (Hymenoptera, Eulophidae), que aparece al final tras los otros dos depredadores y actúa junto con estos.
Palabras clave: Gynaikothrips ficorum, Ficus retusa, Depredadores, Parásito, Argelia
Resum
Fluctuacions en les poblacions de Gynaikothrips ficorum (Marchal, 1908) (Thysanoptera, Tubulifera) i dels seus enemics naturals a les gales de fulles de Ficus retusa a Algèria
L’estudi consisteix en l’observació de les fluctuacions mensuals de les poblacions d’aranyols en Ficus retusa, així com dels seus paràsits i depredadors coneguts, a Boudouaou. Amb aquest objectiu, cada quinze dies es van recollir 20 fulles de cadascun dels arbres seleccionats, durant 12 mesos de gener a desembre de 2015. Aparentment, el nombre d’ous augmenta considerablement a l’agost, quan se’n van registrar 11.163, valor que es va superar lleugerament a l’octubre, amb 11.471 ous. Durant la primavera i l’hivern es va registrar una absència gairebé total d’ous. Se superposen els estadis larvaris 1 i 2 (L1 i L2): L1 va d’agost a octubre, amb 27 i 51 individus respectivament, seguit d’ L2, amb 65 i 186 individus d’octubre a desembre. Les prepupes estan actives a l’abril i al juny, quan assoleixen 46 i 50 individus, respectivament, i augmenten fins a 266 al novembre, si bé mostren valors elevats al juny i al juliol, amb 171 i 135 exemplars respectivament. Els individus adults augmenten progressivament fins a arribar al màxim al començament de desembre, amb 642 individus. També s’han identificat dos depredadors: Montandoniola confusa (Hemiptera, Anthocoridae) i Nephus peyerimhoffi (Coleoptera, Coccinellidae) i només un paràsit: Tetrastichus gentile (Hymenoptera, Eulophidae), que apareix al final després dels altres depredadors i actua juntament amb aquests.
Paraules clau: Gynaikothrips ficorum, Ficus retusa, Depredadors, Paràsit, Algèria
Introduction
About 6,227 species of Thysanoptera are known worldwide (Mound, 2018), 300 species of which produce galls in various plant species (Ananthakrishnan, 1993; Crespi et al., 1997; Lewis, 1997; Mound, 2003). Forty species of the genus Gynaikothrips (Zimmermann) are cecidophagous or galligenous, and originate from Southeast Asia, but have been described from Africa; they are associated with Moraceae plants and especially infest the ornamental Ficus and fruiting figs causing considerable damage (Mound, 2009; Dang et al., 2014). Among these species, Gynaikothrips ficorum (Marchal), also known as Cuban laurel thrips and G. uzeli (Zimmermann), are considered the most studied species due to their similar morphological traits (Priesner, 1939).
G. ficorum (Marchal) is a pantropical species introduced to Algeria from Southeast Asia and first recorded by Del Cañizo (1945). It prefers leaf-gall on Ficus microcarpa L. var. nitida (Urticales, Moraceae) (Priesner, 1939; Alford, 2013; Tree and Wolter, 2009), but in case of abundant infestation, it feeds on other hosts, particularly other genus of Ficus, namely F. axillaris, F. aurea, F. benjamina, F. elastica, F. retusa, as well as Codiaeum variegatum, Melicocca bijuga, Nicotiana tabacum, Viburnum suspensum and Citrus sp. It has also been found on Eucalysptus sp. in Cuba, Gliricidia in Puerto Rico, Calicarpum sp. in Colombia (Denmark, 1967), and occasionally on orchids (Nixon, 1999). Feeding on tender leaves causes the formation of purplish red spots on the underside of the leaves. These become curled and form galls where G. ficorum matting, oviposition and larval development take place (Mound and Marullo, 1996; Morcos, 1944; Wolcott, 1953).
Other pests are also associated with G. ficorum within the leaf galls, such as Chrysomphalus pinnulifer (Diaspididae), Singhiella citrifoli (Aleyrodidae), Greenidea ficicola (Aphididae) (Tawfik, 1967; Paine, 1992; Held et al., 2005; Narrea-Cango et al., 2013; Mifsud et al., 2012; Antonatos et al., 2015). However, several authors have reported the presence of predators on leaf galls: Ontandoniola moraguesi, Orius albidipennis, Macrotracheliella laevis, Cardiastethus rugicollis, Anthochoris flavipes, Anthocoris nemoralis (Hemiptera, Anthocoridae), Termatophylum insigne (Hemiptera, Termatophylidae), Cardiastethus rugicollis, Chrysoperla carnea (Neuroptera, Chrysopidae), and Androthrips ramachandrai (Thysanoptera, Phlaeothripidae). Whereas Adactylidium sp. (Acari), Pleurotropis sp., Tetrastuchus thripophonus, and Tetrastichus gentilei (Hymenoptera, Eulophidae) are considered as parasites of G. ficorum (Dozier (1926); Wolcott (1953); Tawfik (1967); Varadarasan and Ananthakrishnan (1981); Paine (1992); La Salle (1994); Held et al. (2005); Hoddle et al. (2012); Melo et al. (2013); Antonatos et al. (2015))
Several studies on insecticidal control (Wolfenbarger, 1946; Wolcott, 1953; Reinert, 1973) confirm that insecticides can be used to control G. ficorum (Held and Boyd, 2008). Systemic insecticides appear to provide better control than broad-spectrum insecticides that are detrimental to natural enemies that potentially naturally maintain populations of Gynaikothrips at relatively low numbers (Arthurs et al., 2011).
The aims of this study were to study the monthly fluctuation of G. ficorum, to identify their natural enemies, and to explore the impact of these enemies on the population of Ficus retusa in an urban landscape in Algeria.
Material and methods
Study site
This study was conducted in 2015 in a sub-humid area located in the city of Boudouaou (Algeria) at an elevation of about 22 m (36° 43′ 53″ North, 3° 24′ 17″ Est), in the eastern limit of the Mitidja, approximately 35 km from Algiers and 11 km south-west of Boumerdes (fig. 1).

Fig. 1. Map location of sampling area in Boudouaou, Algeria. Fig. 1. Mapa de localización de la zona de muestreo en Boudouaou, Argelia.
This area is characterized by a Mediterranean climate with hot summers and cold winters. The precipitation is irregular; it is abundant in November to March and low from April to September, with a total annual rainfall of 562.4 mm/year (data from the National Meteorology Service, Algiers).
Sampling and analyses of galls
Sampling was carried out randomly on 30 trees. Twenty leaves were collected from each tree (i. e. 600 leaves). Exposure and leaf age were not taken into account. The leaves were placed in plastic bags on which the sampling date, tree number and location were indicated. In the laboratory, the leaf galls were preserved in 70 % ethanol until their mounting and identification. The fortnightly samples were taken from January to December 2015. Specimens were placed in cold KOH 10 % for 24 hours. They were successively bathed in distilled water three times before being mounted in Faure liquid. The counting involved monitoring the demographic evolution of immature stages and adults using a binocular loupe of the type “Carl Zeiss” at 3.6X magnification.
The identification of G. ficorum, including eggs was carried out on the basis of identification keys initiated by Marchal (1908), Del Cañizo (1945), Mound et al. (1976), Quintanilla (1980), Pelikán (1991), Mound et al. (1995), Dang et al. (2014), and Curis et al. (2015).
Statistical analyses
Simple analyses of variance were carried out to verify the period of abundance of each stage and to correlate this with the abundance of their enemies in order to test their impact on the thrips.
Results
Fluctuation of larval stages
The first larval stage (L1) was present all year except from mid-May to mid-August. Activity of L1 and L2 overlapped for two periods. The first period was from February to the end of April and the second was from mid-August to the beginning of October. In the first period, the maximum number of individuals was recorded at the beginning of April for both L1 (114) and L2 (159). In the second period, the maximum was recorded at the end of September and early October for both L1 (67 individuals) and L2 (186 individuals), respectively (fig. 2).
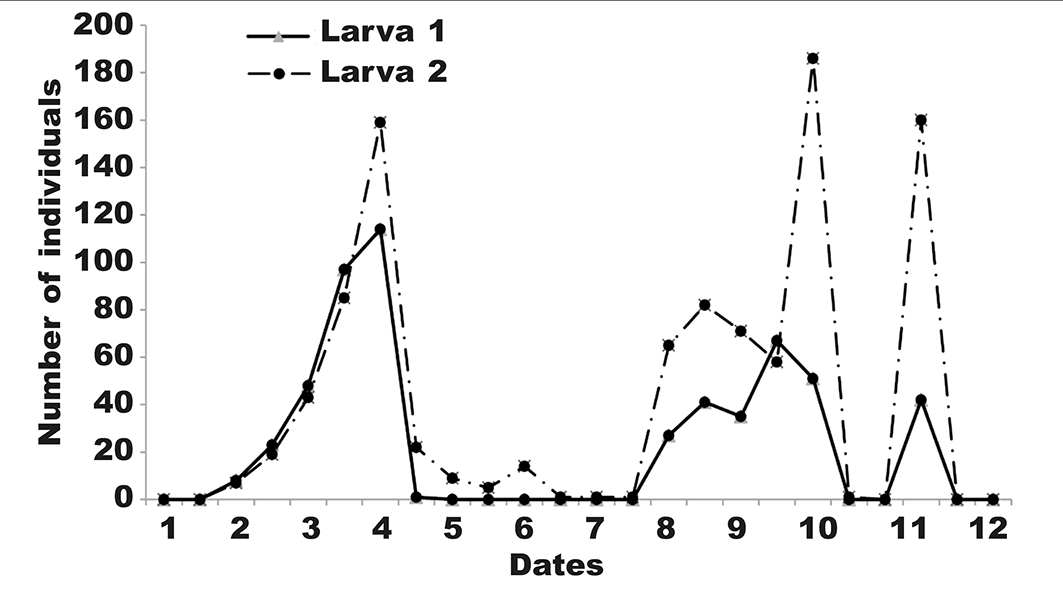
Fig. 2. Monthly evolution of larvae L1 and L2 of G. ficorum on F. retusa in Boudouaou, Algeria. Dates: 1, 01.01.2015; 2, 01.02.2015; 3, 01.03.2015; 4, 05.04.2015; 5, 15.05.2015; 6, 15.06.2015; 7, 15.07.2015; 8, 10.08.2015; 9, 05.09.2015; 10, 01.10.2015; 11,10.11.2015; 12, 01.12.2015. Fig. 2. Evolución mensual de larvas L1 y L2 de G. ficorum en F. retusa en Boudouaou, Argelia. (Para las abreviaturas de las fechas véase arriba).
Fluctuation of pupal stage
The population number of G. ficorum, prepupa and pupa was zero from January to early April and from late June to late July. Numbers increased slightly at the end of April (46 individuals) to reach 50 individuals in mid-June for the prepupae and 171 then 135 individuals for the pupae. Numbers increased again from August to December with highest number at the end of November with 266 individuals for the prepupae and 457 individuals for the pupae (fig. 3).

Fig. 3. Monthly evolution of prepupae and pupae of G. ficorum on F. retusa in Boudouaou, Algeria. Dates: 1, 01.01.2015; 2, 01.02.2015; 3, 01.03.2015; 4, 05.04.2015; 5, 15.05.2015; 6, 15.06.2015; 7, 15.07.2015; 8, 10.08.2015; 9, 05.09.2015; 10, 01.10.2015; 11,10.11.2015; 12, 01.12.2015. Fig. 3. Evolución mensual de prepupas y pupas de G. ficorum en F. retusa en Boudouaou, Argelia. (Para las abreviaturas de las fechas véase arriba).
Fluctuation of G. ficorum adults and eggs
Eggs were absent in January. Their number gradually increased to 4,280 eggs in early April then declined significantly in mid-May to 491 eggs and in mid-July to 145. The number of eggs later increased, and was highest in early October (11,471 eggs). Numbers then fell to 6,454 in November, but increased again in mid-December to 10,400 eggs.
The adult population was present almost year-round, with the exception of early February to early April when the population was negligible. Adults appeared from the end of April, with 117 individuals, and continued to increase in the following months to reach a peak in early December with 642 individuals (fig. 4).

Fig. 4. Monthly evolution of eggs and adults of G. ficorum on F. retusa in Boudouaou, Algeria. Dates: 1, 01.01.2015; 2, 01.02.2015; 3, 01.03.2015; 4, 05.04.2015; 5, 15.05.2015; 6, 15.06.2015; 7, 15.07.2015; 8, 10.08.2015; 9, 05.09.2015; 10, 01.10.2015; 11,10.11.2015; 12, 01.12.2015. Fig. 4. Evolución mensual de huevos y adultos de G. ficorum en F. retusa en Boudouaou, Argelia. (Para las abreviaturas de las fechas véase arriba).
Damage of G. ficorum on F. retusa
G. ficorum adults fed on young leaves and induced the appearance of purplish red spots along the main vein, causing curling of the leaf. Infested leaves dehydrated, hardened and changed color gradually, turning from yellow to brown before falling out on windy or rainy days. This damage did not generally kill the tree but did decrease their ornamental value (fig. 5).

Fig. 5. The G. ficorum damages on the leaves of Ficus retusa in different stage of maturation. Fig. 5. Daños causados por G. ficorum en las hojas de Ficus retusa en distintos estadios de maduración.
Predators and parasites of the G. ficorum
Gall testing revealed important diversity of associated enemies within the gall leaf. We found three major enemies of G. ficorum that were present in all the life cycle of this species.
Montandoniola confusa (Streito and Matocq, 2009) (Hemiptera, Anthocoridae)
We found this predator to be present in high numbers in summer, reaching 50 individuals in August. This parasite feeds on all stages of G. ficorum: eggs, larvae, prepupae, and pupae (fig. 6).

Fig. 6. Monthly evolution of M. confusa on Ficus retusa in Boudouaou, Algeria. Dates: 1, 01.01.2015; 2, 01.02.2015; 3, 01.03.2015; 4, 30.04.2015; 5, 15.05.2015; 6, 15.06.2015; 7, 15.07.2015; 8, 10.08.2015; 9, 05.09.2015; 10, 01.10.2015; 11,10.11.2015; 12, 01.12.2015. Fig. 6. Evolución mensual de M. confusa en Ficus retusa en Boudouaou, Argelia. (Para las abreviaturas de las fechas ver arriba.)
Nephus peyerimhoffi (Sicard, 1923) Oenopia doublieri (Mulsant, 1846)
N. peyerimhoffi (Coleoptera, Coccinellidae) is coccidiphagous, highly present in leaf galls, particularly in the spring when its main food sources Pseudococcus species, i.e. Pseudococcus citri (Risso) and Pseudococcus adonidum (Linné) (Saharaoui et al., 2015) are present. Few individuals of Oenopia doublieri were observed during our sampling. N. peyerimhoffi is a predatory thrip that feeds mainly on eggs, but can also consume larval and pupal stages; it was present throughout the study period and its presence was essential during the period of thrip laying. Abundance was significant in February (55 individuals) (fig. 7).

Fig. 7. Monthly evolution of N. peyerimhoffi on Ficus retusa in Boudouaou, Algeria. Dates: 1, 01.01.2015; 2, 01.02.2015; 3, 01.03.2015; 4, 05.04.2015; 5, 15.05.2015; 6, 15.06.2015; 7, 15.07.2015; 8, 10.08.2015; 9, 05.09.2015; 10, 01.10.2015; 11,10.11.2015; 12, 01.12.2015. Fig. 7. Evolución mensual de N. peyerimhoffi en Ficus retusa en Boudouaou, Argelia. (Para las abreviaturas de las fechas ver arriba.)
Tetrastichus gentilei (Del Guercio, 1931)
T. gentilei (Hymenoptera, Eulophidae) is an endoparasite of larval stages of thrips (fig. 8). During the surveys it was observed from August to December. Its population varied from 20 to 34 individuals in August, then dropped in November before starting increasing slightly again to four and 10 individuals respectively in early and late December.

Fig. 8. Monthly evolution of Tetrastichus gentilei on Ficus retusa in Boudouaou, Algeria. Dates: 1, 01.01.2015; 2, 01.02.2015; 3, 01.03.2015; 4, 05.04.2015; 5, 15.05.2015; 6, 15.06.2015; 7, 15.07.2015; 8, 10.08.2015; 9, 05.09.2015; 10, 01.10.2015; 11,10.11.2015; 12, 01.12.2015. Fig. 8. Evolución mensual de Tetrastichus gentilei en Ficus retusa en Boudouaou, Argelia. (Para las abreviaturas de las fechas ver arriba.)
Succession of presence of M. confusa, N. peyerimhoffi and T. gentilei
The frequency of the predators and parasites overlapped among the species.The ladybird N. peyerimhoffi was present first, in March, at the time of the appearance of the eggs on which it feeds. It remained throughout the year. It was followed in number by M. confusa, which feeds preferably on eggs, but can consume other larval stages. The parasite T. gentilei was observed in mid-July. M. confuse and N. peyerimhoffi were present in late June. T. gentilei appeared at the beginning of August when the predators and the parasites acted together (fig. 9). Based on our observations, M. confusa was the most abundant insect in the analyzed leaf galls. It was considered the major predator of several species of thrips.
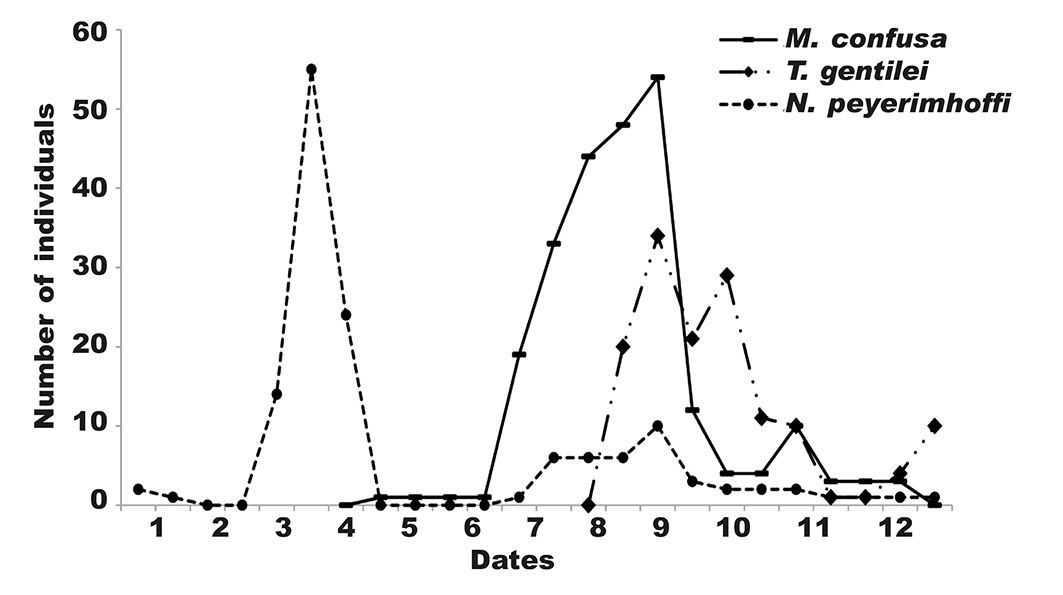
Fig. 9. Ordre of succession of predators and parasite of G. ficorum on F. retusa in Boudouaou, Algeria. Dates: 1, 01.01.2015; 2, 01.02.2015; 3, 01.03.2015; 4, 05.04.2015; 5, 15.05.2015; 6, 15.06.2015; 7, 15.07.2015; 8, 10.08.2015; 9, 05.09.2015; 10, 01.10.2015; 11,10.11.2015; 12, 01.12.2015. Fig. 9. Orden de sucesión de predadores y parásitos de G. ficorum en F. retusa en Boudouaou, Argelia. (Para las abreviaturas de las fechas ver arriba.)
Discussion
Data from this study indicate great diversity at all stages of G. ficorum on the gall leaf of Ficus retusa. The number of eggs was particularly diverse and highly significant in August. For the larval stages, abundance was highest in autumn and spring, while for the pupal stage, it was highest in November, likely due to the age of galls and the influence of climatic conditions. However, the population of this species was highest in August, followed by November and October. The high population of G. ficorum in August may be due to the shorter life cycle of thrips and greater opportunity of an overlapping generation (Mascarenhas and Silva, 2016). The presence of the associated predator and parasites on the same gall leaf may impact the population of G. ficorum. Other studies consider that the date of planting may be a factor of variation in the number of thrips; Frankliniella fusca swarm in the early plantings of peanuts in April, while this number decreases in late plantings in May and June (Todd et al., 1995).
According to Paine (1992), the generation of G. ficorum peaks within mature galls during summer, likely due to the high temperature and increase in rainfall. He noted that the population of G. ficorum ranged from 48 to 99 individuals at 15 °C and about 16 individuals at 13 °C, and also suggested that lower and upper temperature thresholds were observed at 12 and 35 °C, suggesting that spring temperatures do not limit the development of thrips. However, Arthurs et al. (2011) suggested that the temperature variation has an impact on the cycle life of the genus Gynaikothrips under greenhouses conditions. Shogren and Pain (2015) mentioned that another factor that may influence the population of thrips is the sampling period. They found the percentage of thrips differed at the same study site over the different sampled years, resulting in an abundance of thrips within the galls. However, Rivnay (1974) found that the population of G. ficorum increased from March to June, and also in late August and October, but decreased in mid-summer. This is comparable to our results and may be due to more abundant and diverse fauna at these times.
Among specimens of the gall-associated fauna, we highlighted two major predators, M. confusa and N. peyerimhoffi, previously associated with G. ficorum and T. gentilei as a parasite of G. ficorum. Generally, the species of the genus Montandoniola are predators of most thrips (Dobbs and Boyd, 2006; Yamada et al., 2011). In this study, G. ficorum predators were present in greater quantity, particularly M. confuse. This led to a sharp decrease in numbers of this predator. It also impacted on the mortality of eggs, and decreased the population from 11,471 eggs to about 200 larvae. On this subject, Arthurs et al. (2011) noted that M. confusa can reduce the population of G. ficorum by 95 % and leaf galls by up to 7 7 % within 5 weeks. In a study on G. ficorum, Funasaki (1966) reported that the nymphs in the 1st and 2nd instars of M. moraguesi feed on eggs and larvae of G. ficorum, and nymphs in the 3rd, 4th and 5th instars feed on thrips at any stage of development, including adults. Tavares et al. (2013) mentioned that M. confusa feeds on all stages of G. ficorum, but it prefers the eggs, likely due to their nutritional quality and their availability compared to the other stages. Regarding the parasites of G. ficorum, the females of T. gentilei found in this investigation laid their eggs in the young larvae of G. ficorum, presumably the prepupae, pupae and the adults, including the larvae. Bournier (1967) found that the second larval stages L2 were never attacked by this parasite, and our results confirmed this. Boyd and Held (2016) found that wasps can be used as a parasitoid of the 1st and 2nd instars of Gynaikothrips uzeli with an emergence time twice as fast as on G. ficorum. It was also revealed that
T. gentilei is a parasitoid of guava thrips Liothrips anonae (Pikart et al., 2015).
In conclusion, we found that the abundance of G. ficorum was highest in August and decreased significantly in winter, likely due to the effect of their associated natural enemies, particularly, M. confusa, that strongly reduced the populations of G. ficorum. We also noted a higher opportunity for overlap among generations in 2015 and synchronization between this generation and the presence of their enemies. The interaction of Thrip-enemies is a complex factor that requires further study to understand the impact of the associated predators and parasites on the population of thrips in Algeria.
Acknowledgements
We would like to thank my supervisor Abdelmadjid Benzehra for support and encouragement, and Hezil Sara and Manar Benoumhani for help with the English language.
















