
Arxius de Miscel·lània Zoològica. Volumen 19 (2021) Páginas: 83-98
First genetic identification of Sundevall's jird Meriones crassus (Rodentia, Muridae) in Morocco and the hypothesis of range extension
Bouarakia, O., Benhoussa, A., Lalis, A., Benazzou, T., Bonillo, C., Denys, C.
DOI: https://doi.org/10.32800/amz.2021.19.0083Descargar
PDFCita
Bouarakia, O., Benhoussa, A., Lalis, A., Benazzou, T., Bonillo, C., Denys, C., 2021. First genetic identification of Sundevall's jird Meriones crassus (Rodentia, Muridae) in Morocco and the hypothesis of range extension. Arxius de Miscel·lània Zoològica, 19: 83-98, DOI: https://doi.org/10.32800/amz.2021.19.0083-
Fecha de recepción:
- 07/12/2020
-
Fecha de aceptación:
- 17/03/2021
-
Fecha de publicación:
- 20/04/2021
-
Compartir
-


-
Visitas
- 4461
-
Descargas
- 1270
Abstract
First genetic identification of Sundevall’s jird Meriones crassus (Rodentia, Muridae) in Morocco and the hypothesis of range extension
Rodents from the subfamily Gerbillinae Gray, 1825, are an important component of small fauna in arid and desertic regions in Africa and Asia. Among this group of rodents, Sundevall’s jird, Meriones crassus, is one of the most widespread desert species, found in countries ranging from Morocco to Pakistan. In this study, we sequenced the cytochrome b gene of the mitochondrial DNA in two specimens captured in Morocco, constituting the first genetic identification for this species in the African continent. Molecular identification was complemented and supported with morphological evidence. Additionally, we propose the hypothesis of a northward extension of 250 km of the geographic distribution of Sundevall’s jird in Morocco, perhaps indicating the increasing desertification affecting North Africa. Further evidence will be required to test the latter hypothesis.
Dataset published through Zenodo (Doi: 10.5281/zenodo.4674889)
Key words: Distribution, Mitochondrial DNA, North Africa, Desert, Morphology, Phylogenetics
Resumen
Primera identificación genética del jerbo de Sundevall Meriones crassus (Rodentia, Muridae) en Marruecos y la hipótesis de extensión de su ámbito de distribución
Los roedores de la subfamilia Gerbillinae Gray, 1825, representan un importante componente de la fauna de pequeño tamaño de regiones áridas y desérticas de África y Asia. El jerbo de Sundevall Meriones crassus es una de las especies desérticas más extendidas de esta subfamilia y está distribuido desde Marruecos hasta Pakistán. En este estudio secuenciamos el gen citocromo b del ADN mitocondrial de dos especímenes capturados en Marruecos, lo que constituye la primera identificación genética de esta especie en el continente africano. La identificación molecular se complementó y confirmó con evidencia morfológica. Además, propusimos la hipótesis de una extensión de su distribución geográfica 250 km hacia el norte en Marruecos, quizá indicativa de la creciente desertificación que afecta al norte de África. La confirmación de dicha hipótesis requerirá pruebas adicionales.
Datos publicados en Zenodo (Doi: 10.5281/zenodo.4674889)
Palabras claves: Distribución, ADN mitocondrial, Norte de África, Desierto, Morfología, Filogenético
Resum
Primera identificació genètica del jerbu de Sundevall Meriones crassus (Rodentia, Muridae) al Marroc i la hipòtesi d’extensió del seu àmbit de distribució
Els rosegadors de la subfamília Gerbillinae Gray, 1825, representen un component important de la fauna de mida petita de regions àrides i desèrtiques d’Àfrica i Àsia. El jerbu de Sundevall Meriones crassus és una de les espècies desèrtiques més esteses d’aquesta subfamília i està distribuït des del Marroc fins al Pakistan. En aquest estudi seqüenciem el gen citocrom b de l’ADN mitocondrial de dos espècimens capturats al Marroc, la qual cosa constitueix la primera identificació genètica d’aquesta espècie al continent africà. La identificació molecular es va complementar i confirmar amb evidència morfològica. A més a més, vam proposar la hipòtesi d’una extensió de la seva distribució geogràfica 250 km cap al nord al Marroc, potser indicativa de la creixent desertització que afecta el nord d’Àfrica. La confirmació d’aquesta hipòtesi requerirà proves addicionals.
Dades publicades a Zenodo (Doi: 10.5281/zenodo.4674889)
Paraules clau: Distribució, ADN mitocondrial, Nord d’Àfrica, Desert, Morfologia, Filogenètic
Introduction
Jirds from the genus Meriones Illiger, 1811 are a group of relatively large and heavily-built Gerbillinae rodents distributed from North Africa to Mongolia. This genus comprises 16 species, but only three species are generally recognized in the African continent: Sundevall’s jird Meriones crassus Sundevall, 1842; the Libyan jird M. libycus Lichtenstein, 1823; and Shaw’s jird M. shawii Duvernoy, 1842 (Fichet-Calvet, 2013). This latter taxon is structured as a species complex and is separated by some authors into two distinct species: M. shawii and M. grandis Cabrera, 1907 (Denys et al., 2017). This species complex will be hereafter referred to as M. shawii sensu lato. Among these three to four aforementioned North African species, M. crassus is the truly desertic one. It occupies the sandy or hammada desert environments from the Atlantic Sahara to Pakistan, including the Middle East and the Arabian Peninsula. In Morocco, it inhabits the southernmost regions in the south of the Anti-Atlas and of the High-Atlas Mountains, and certainly all the Atlantic Sahara (Petter, 1961; Aulagnier et al., 2017).
M. crassus is sympatric with M. libycus in a large part of its geographic and climatic range in Morocco, but M. crassus lives in the most arid habitats that are devoid of vegetation, including sand dunes but excluding rocks. In contrast, M. libycus occupies habitats with more vegetation cover, such as oases, unflooded riverbanks and stabilized dunes (Petter, 1961; Granjon, 2016). On the other hand, M. shawii is found mainly in areas with a Mediterranean climate in the North and in agroecosystems, but a sympatry with M. crassus is possible in the southernmost margins of the distribution of M. shawii (Aulagnier et al., 2017).
In north-eastern Morocco, M. crassus is replaced by M. libycus and M. shawii. This region of Morocco is mainly located on a high plateau, characterized by an arid to semi-arid climate, and partially covered by the basin of the Moulouya, the largest permanent river in Morocco. This region is bordered in the north by the Tell Atlas Mountains and the Mediterranean Sea; in the south by the Sahara Desert; in the west by the Middle Atlas and the Rif Mountains, and in the east by the Algerian High Plateaux and the Saharan Atlas Mountains (Chichagov, 2008). These characteristics make north-eastern Morocco a transitional zone from Mediterranean to desert ecoclimatic regions (Oliva et al., 2018). Despite the importance of this region, at the ecological and zoogeographic levels, knowledge remains incomplete concerning the distribution, ecology and systematics of the small mammals therein.
The study of this fauna first needs proper identification of the different taxa composing it. The identification of Meriones and other Gerbillinae species, based solely on external morphological features cannot be trusted due to the morphological resemblance between numerous species, the high intraspecific variability, and the existence of cryptic species. Thus, the use of other methods such as cranial morphometry, karyotyping or DNA barcoding are required to ensure reliable species identification (Benazzou et al., 1982; Ndiaye et al., 2012, 2013). In Morocco, despite slight variations in external body characteristics among Meriones species (i.e. tail length, body size, dorsal coat coloration, brush at the end of the tail, colour of claws on hind feet) (Darvish, 2011; Denys et al., 2017), they can easily be misidentified, especially in the case of juvenile individuals.
On a cytogenetic level, M. crassus can be easily distinguished from M. libycus (2n = 44, FN = 74) and M. shawii (2n = 44, FN = 78) (Lay and Nadler, 1969; Benazzou et al., 1982; Gerbault-Seureau et al., 2015). The karyotype of M. crassus has been specified to have a stable diploid number of chromosomes (2n = 60) and a fundamental number showing differences between populations from Africa and Asia: 76 in Turkey (Yiğit et al., 1998); 74 in Iran, Syria, Algeria, Jordan and Morocco (Matthey, 1957; Qumsiyeh et al., 1986; Benazzou et al., 1982); 72 in Iran and Egypt (Nadler and Lay, 1967).
Recent phylogeographic studies have provided the first molecular identification of different Gerbillinae and Meriones species in Morocco (Ndiaye et al., 2012; Nicolas et al., 2014; Lalis et al., 2016; Bouarakia et al., 2018, 2019a; Dianat et al., 2020), but the molecular identification of M. crassus has not yet been performed in Morocco. Furthermore, no specimens have been identified using molecular tools in the African continent.
The present study aims to provide the first molecular characterization of Sundevall’s jird M. crassus in Morocco, to describe its morphological characterization, and to review its geographic range in this country.
Material and methods
Sampling in the study area
Field investigation in arid areas of north-eastern Morocco in October-November 2014 allowed us to catch two individuals of M. crassus. The first individual (BRF9) was caught using a Sherman trap on a sandy steppe with Chenopodiaceae shrubs, west of the city of Bouarfa (32° 30′ N, 2° 02′ W), after a trapping effort of 186 trap-nights. The second individual (BMT5) was captured by hand in its burrow in a rocky plateau with Chenopodiaceae shrubs, north of Ain Beni Mathar town (34° 07′ N, 2° 02′ W) (fig. 1).
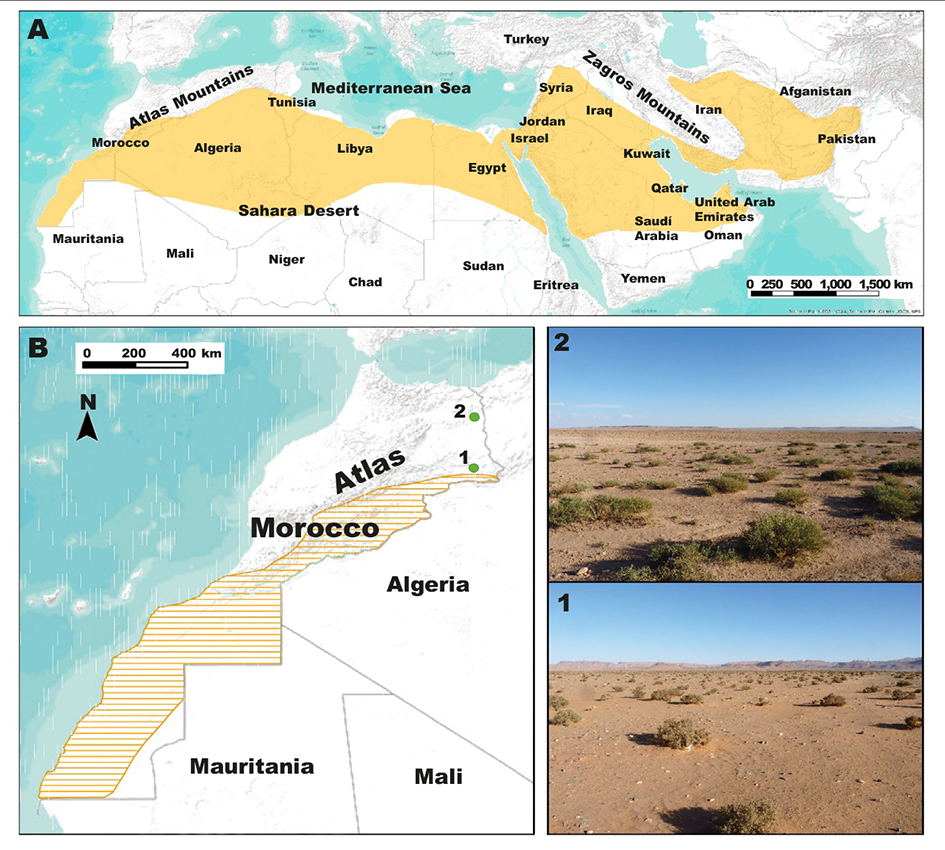
Fig. 1. A, entire known distribution range of Meriones crassus; B, distribution range of Meriones crassus in Morocco based on confirmed observations from the literature and the two new sampled localities: Bouarfa (1) and Ain Beni Mathar (2). Fig. 1. A, totalidad del ámbito de distribución conocido de Meriones crassus; B, ámbito de distribución de Meriones crassus en Marruecos basado en observaciones confirmadas procedentes de la literatura y las dos nuevas localidades donde se efectuaron capturas: Bouarfa (1) y Ain Beni Mathar (2).
Prior to the study, all necessary permits were obtained; we complied with the pertinent protection regulations in force and followed all applicable Moroccan and institutional guidelines for the care and use of animals. The two animals were euthanized by cervical dislocation. This protocol was approved by the Cuvier Ethics Committee (National Museum of Natural History of Paris, France).
Morphological study
Morphological identification was initially based on external body features (e.g., back coat, colour of hind feet claws, brush at the end of the tail) (fig. 2). The two samples were then weighed (WT) (in g) and head-body length (HB), tail length (T), hind feet length (HF) and ear length (E) were calculated in mm. We also calculated the ratio of tail length to head-body length (%T).

Fig. 2. Young adult male of Meriones crassus (BRF9). Fig. 2. Ejemplar de macho adulto joven de Meriones crassus capturado (BRF9).
The skull was extracted (fig. 3) and features were described. We took eight measurements (in mm) on dorsal and ventral sides of the skull using a Mitutoyo calliper that was accurate to 0.01 mm: greatest length of skull (GLS), breadth of braincase (BB), least interorbital constriction (IO), length of nasals (LN), width of zygomatic arch (WZYG), length of anterior palatine foramina (LAF), length of upper molar series (M1M3) and diagonal length of tympanic bulla (LTB). We also calculated the ratio of tympanic bulla length to skull length (%TB).

Fig. 3. Skulls of the individuals BRF9 and BMT5 on the dorsal (d) and ventral side (v). Fig. 3. Vista dorsal (d) y ventral (v) de los cráneos de los individuos BRF9 y BMT5.
The vouchers of the two specimens are stored in the collections of the Faculty of Sciences of Rabat (Mohammed V University in Rabat, Morocco) (‘FSR-MAR14-BRF9’ and ‘FSR-MAR14-BMT5’). For the sake of comparison, we also examined the external body and skull features of genetically confirmed Meriones specimens from Morocco (vouchers ‘FSR-MAR14-TAT1’ for M. libycus and ‘FSR-MAR15-GCF9’ for M. shawii in the collections of the Faculty of Sciences of Rabat).
Molecular study
For the genetic identification, we used the cytochrome b mitochondrial gene, cytb, (1,040 base pairs, bp), the most commonly used DNA marker for delimiting species boundaries in rodents. We extracted the DNA from ethanol-preserved liver using the QIAGEN Kit (DNeasy Blod and Tissue Kit) following the manufacturer recommendations. Then we amplified the cytb gene via polymerase chain reaction using the primers L7 (5′-ACC AAT GAC ATG AAA AAT CAT CGTT-3′) and H6 (5′-TCT TCA TTT TTG GTT TAC AAG AC-3′) (Mongelard et al., 2002). Double-stranded PCR products were purified and sequenced in both directions by Eurofins (France).
We corrected and aligned the two obtained sequences both manually and using BioEdit version 7.2 (Hall, 1999). The two partial sequences (788 bp for BRF9 and 748 bp for BMT5) were submitted to GenBank database (see table 1). We included in the genetic analysis all the cytb sequences for M. crassus present in GenBank (three in total). We also included representatives of the other species of the genus Meriones found in GenBank (10 species). We aligned all these sequences and maintained a fragment of 712 bp for the analysis in view of the limited length of one M. crassus sequence in the GenBank.

Table 1. Cytochrome b sequences used in the phylogenetic analysis. Tabla 1. Secuencias de citocromo b utilizadas en el análisis filogenético.
We estimated the evolutionary relationships between the sequences and constructed the phylogenetic tree based on the maximum likelihood analysis (ML) in MEGA version 7 (Tamura et al., 2013) and the Bayesian inference analysis (Bayesian Markov Chain Monte Carlo approach, MCMC) using MrBayes version 3.2 (Ronquist et al., 2012). For these analyses, we used MrModeltest version 2.4 (Nylander, 2004) to determine the general time reversible (GTR) + I + G model (Gu et al., 1995) as the best-fit model of nucleotide substitution, according to the Akaike information criterion (Akaike, 1973). The phylogenetic analyses were performed on a non-partitioned alignment.
In the maximum likelihood analysis, the robustness of the obtained topologies was tested in all the treatments using 1,000 bootstrap replicates, and the BIONJ distance-based tree was used as the starting tree. Bayesian posterior probabilities (PP) were used to measure statistical support in the Bayesian inference analysis. In this latter analysis, two independent MCMC runs of five million generations, with burn-ins of 25 %, were conducted and the trees and parameters were sampled every 100 generations. Stationarity was then evaluated by examining the average standard deviation of split frequencies and the potential scale reduction factor (Ronquist et al., 2012).
Two species from the Gerbillinae subfamily were used as outgroups in the phylogenetic tree (Psammomys obesus Cretzschmar, 1828 and Gerbillus simoni Lataste, 1881). Additionally, we calculated the pairwise Kimura two-parameter (K2P) genetic distances (Kimura, 1980) between sequences using MEGA 7.
Results
Morphological identification
Table 2 shows the body and cranial measurements of the two individuals BRF9 (young adult male) and BMT5 (juvenile male) (see also the dataset published through Zenodo, Doi: 10.5281/zenodo.4674889).

Table 2. Body and cranial measurements of the two collected individuals and comparison with measurements of Meriones crassus from the literature: I, individuals or reference; WT, weight in g; HB, head–body length; T, tail length; %T, ratio of tail length to head–body length; HF, hind feet length; E, ear length; GLS, greatest length of skull; BB, breadth of braincase; IO, least interorbital constriction; LN, length of nasals; WZYG, width of zygomatic arch; LAF, length of anterior palatine foramina; M1M3, length of upper molar series; LTB, diagonal length of tympanic bulla; %TB, ratio of tympanic bulla length to skull length; min, minimum; max, maximum. (Measurements in mm.) Tabla 2. Medidas corporales y craneales de los dos individuos capturados y comparación con las medidas de Meriones crassus registradas en la literatura: I, individuos o referencia; WT, peso en g; HB, longitud cabeza–cuerpo; T, longitud de la cola; %T, relación entre la longitud de la cola y la longitud cabeza–cuerpo; HF, longitud de las extremidades posteriores; E, longitud de la oreja; GLS, diámetro mayor del cráneo; BB, ancho de la cavidad craneal; IO, menor constricción interorbital; LN, longitud de los nasales; WZYG, ancho del arco cigomático; LAF, longitud del foramen palatino anterior; M1M3, longitud de la serie molar superior; LTB, longitud diagonal de la ampolla timpánica; %TB, relación entre la longitud de la ampolla timpánica y la longitud del cráneo; mín, mínimo; máx, máximo. (Medidas en mm.)
Measurements of both individuals fit within the measurements of M. crassus found in the literature (table 2). Several M. crassus measurements can be differentiated from those of the other two Moroccan Meriones species, especially weight (55-158 g in M. crassus, 60-140 g in M. libycus, 230-255 g in M. shawii), the ratio of tail length to head-body length (92-109 % in M. crassus, 107-113 % in M. libycus, 90-99 % in M. shawii), and the ratio of tympanic bulla length to skull length (35-41 % in M. crassus, 33-39 % in M. libycus, 28-31% in M. shawii) (Petter, 1961; Ranck, 1968; Osborn and Helmy, 1980; Aulagnier et al., 2017). The external body and cranial features of both individuals are consistent with descriptions of M. crassus (Petter, 1961; Harrison and Bates, 1991; Kowalski and Rzebik-Kowalska, 1991; Darvish, 2011; Tabatabaei Yazdi et al., 2014; Aulagnier et al., 2017; Denys et al., 2017). They exhibit a soft, fawn-coloured back coat. The tail is relatively short (the same length as the head-body or slightly shorter than the head-body) with a small terminal brush of dark hair. The hind feet claws are whitish. The tympanic bullae are hypertrophied, extending far beyond the occipital condyle. The swelling of the auditory meatus lip is very conspicuous and is in contact with the zygomatic arch. The suprameatal triangle of the tympanic bulla is large and open at its posterior end (fig. 3).
Genetic identification
The maximum likelihood and Bayesian inference phylogenetic analyses produced similar topologies, especially regarding the clade containing the sequences from M. crassus and its two closest congener species (the king jird Meriones rex Yerbury and Thomas, 1895 and Tristram’s jird Meriones tristrami Thomas, 1892). For the sake of simplicity, we present only the phylogenetic tree originating from the maximum likelihood analysis in figure 4, but specify Bayesian posterior probabilities. Although the phylogenetic resolution between species is low across most of the tree, the two Moroccan individuals (BRF9 and BMT5) from the localities of Bouarfa and Ain Beni Mathar show low levels of genetic differentiation. Most importantly, they are nested unambiguously with the specimens of M. crassus with high bootstrap values (≥ 91 %) and high posterior probabilities (≥ 0.9). The Moroccan individuals and the three M. crassus specimens from the Middle East (Sinai Peninsula, Saudi Arabia and Iran) form a strongly supported monophyletic group (bootstrap value of 99 % and posterior probabilities of 0.99), supporting the identification of these individuals as M. crassus.

Fig. 4. Maximum likelihood phylogenetic tree based on the cytochrome b gene, depicting the relationships between the two Meriones crassus specimens (BFR9 and BMT5), other conspecific populations, and Meriones species. Psammomys obesus and Gerbillus simoni were used as outgroups. Values at nodes represent ML bootstrap support (given only if ≥ 50 %) and Bayesian PP (given only if ≥ 0.9). The scale bar represents the branch length measured in the number of substitutions per site. The geographic origin of M. crassus specimens is indicated on the right–hand side of the figure. Fig. 4. Árbol filogenético de máxima verosimilitud basado en el gen citocromo b, que representa las relaciones entre los dos especímenes de Meriones crassus (BFR9 y BMT5), otras poblaciones coespecíficas y especies de Meriones. Psammomys obesus y Gerbillus simoni fueron utilizados como grupos externos. Los valores en los nodos representan el soporte de bootstrap ML (dado solo si ≥ 50 %) y PP bayesiano (dado solo si ≥ 0,9). La barra de escala representa la longitud de rama medida en el número de sustituciones por emplazamiento. El origen geográfico de los especímenes de M. crassus se indica en el lado derecho de la figura.
Within the M. crassus group, the Moroccan individuals are genetically closer to the specimens from the Sinai Peninsula and Saudi Arabia (2.3 % K2P) than to the Iranian specimen (3.9 % K2P). The closest species to M. crassus are M. rex (5.8 % K2P) and M. tristrami (7.6 % K2P).
Discussion
External morphological differentiation between Moroccan Meriones species (M. crassus, M. libycus and M. shawii) is slight. For example, the tail is relatively longer in M. libycus, the body is heavier in M. shawii, the dorsal coat is a lighter colour in M. crassus, the hind feet claws are blackish in M. libycus, and the brush at the end of the tail is more developed in M. libycus. As such differences are small, identification is particularly challenging in juvenile individuals (Darvish, 2011; Denys et al., 2017).
Examination of discrete morphological features and body/craniodental morphometric measurements allowed us to identify the two individuals as M. crassus. The tympanic bulla of M. crassus is generally the longest among the Moroccan Meriones species, and its mastoid bulla is very large and extends well posterior to the para-occipital process compared to M. libycus whose tympanic bulla extends slightly posterior to the para-occipital process and to M. shawii whose tympanic bulla just reaches the para-occipital process. In addition, the swelling of the auditory meatus lip is very visible and is always in contact with the zygomatic arch in M. crassus. In comparison, in M. libycus, this swelling is visible but to a lesser degree and it is usually in contact with the zygomatic arch, while in M. shawii, this swelling is just slightly visible and it never touches the zygomatic arch. In M. crassus, the suprameatal triangle is wide open and joins the tympanic bulla, while it is partially open with a narrow opening to the tympanic bulla in M. shawii, and it is closed and does not join the tympanic bulla in M. libycus (Petter, 1961; Ranck, 1968; Osborn and Helmy, 1980; Fichet-Calvet, 2013).
Since M. libycus is the closest congener and sympatric with M. crassus in Morocco, it is essential to identify the diagnostic characters of both species as tympanic bullae dimensions and dental characteristics may overlap and confuse identification. It is also interesting to note that in the literature, M. crassus longifrons Lataste, 1884 and M. crassus charon Thomas, 1919, two subspecies of M. crassus, possess less swollen bullae than the other subspecies described within this species. Moreover, M. libycus arimalius Cheesman and Hinton, 1924, one of the subspecies of M. libycus, is defined as having the largest tympanic bullae of the species, making them closer in size to those of M. crassus (Ellerman, 1948; Wittmer and Büttiker, 1982).
Additionally, the inflation of the tympanic bullae increases significantly, on an intraspecific level, in Meriones and Gerbillinae species inhabiting arid or desertic areas (Alhajeri et al., 2015; Bouarakia et al., 2019b). A two-dimensional geometric analysis of M. crassus skulls originating from the geographic range of the species, but mostly those from the Middle East, showed cranial differences between populations, mainly at the level of the relative size of the tympanic bulla. These differences were significantly correlated with the annual rainfall and elevation (Tabatabaei Yazdi et al., 2014). A geometric morphometric study including more samples from Africa and the whole geographic range of M. crassus is necessary to better evaluate the extent of its intraspecific morphological diversity and to provide a more accurate distinction from the other closely related jirds.
Phylogenetic evidence based on the cytochrome b gene supported our morphological identification of the two individuals, representing the first genetic characterization of M. crassus in Morocco and the African continent. Although sampling is very limited, our genetic analysis reveals the existence of some genetic structure within the geographic range of this species, and a higher sequence divergence between the most distant localities (Morocco and Iran). The intraspecific divergence shown across the wide range of this Saharan species is probably due to the great geographic distance existing between the populations. An uneven occurrence of populations throughout the range of the species and the presence of biogeographic barriers to gene flow (e.g., Nile River in Egypt, Zagros Mountains in Iran) could also play a role in this genetic structuration. Previous studies have reconstructed the phylogeography of Gerbillines inhabiting Morocco, including Gerbillus species (Ndiaye et al., 2012; Nicolas et al., 2014; Bouarakia et al., 2018, 2019a; 2019b), as well as the M. shawii species complex and M. libycus (Lalis et al., 2016; Khemiri et al., 2017; Dianat et al., 2020). Subsequently, identification of the existing lineages of M. crassus and a detailed description of its phylogeographic structuration at the scale of North Africa are still pending.
Various subspecies of M. crassus are described in the literature (e.g. M. crassus crassus Sundevall, 1842 from North Africa; M. c. longifrons from the Arabian Peninsula; M. c. charon from the western parts of Middle East; M. c. swinhoei Scully, 1881 from the eastern parts of Middle East) based on geographic distribution and a few body or skull characteristics such as pelage colour, tail length, and the size of the tympanic bullae (Ellerman, 1948; Petter, 1961; Darvish, 2011; Tabatabaei Yazdi et al., 2014). However, the legitimacy of these subspecies, their geographic range and boundaries need to be verified using molecular phylogenies. Sampling from Morocco and across the geographic range of M. crassus in North Africa and Asia is necessary for future studies investigating this widespread jird species.
M. crassus is known in Morocco only from arid/desert areas of the south and south-east of the country, and the previous northernmost record being near the city of Figuig (32° 06′ N, 1° 14′ W), located at the western end of the Saharan Atlas Mountain Range. The individuals captured in the two localities of north-eastern Morocco suggest a northward extension of 250 km of the known species geographic range in Morocco. In Algeria, the northernmost record of the species is located south of the Saharan Atlas Mountain Range in the north-west of the country (Kowalski and Rzebik-Kowalska, 1991). According to Petter (1961), M. crassus in the Sahara could be considered an ecological indicator of the desert environment due to the fact that its limit of distribution towards the north seems to coincide with that of the date palm groves.
The finding presented in our study could be interpreted as a result of the desertification process taking place in North Africa, with the threat of sand advancing towards the northern regions characterised typically by a Mediterranean climate. In northern Algeria, the high plateaux semi-arid zone, a natural region consisting of steppe-like alluvial plains situated north of the Saharan Atlas mountain range, is gradually turning into a desert (Kasbadji Merzouk et al., 2003). Climate change and the increasing aridification witnessed in North Africa can induce an expansion of the Palearctic desert and pre-desert fauna. Similar to the case of M. crassus, the pygmy gerbil Gerbillus henleyi de Winton, 1903, a Gerbillinae widespread in desert and semi-desert regions of North Africa, was also recently discovered in north-eastern Morocco (Bouarakia et al., 2018). Supplementary field surveys are needed to shed more light on these climate-induced expansions. An alternative hypothesis is that M. crassus could have been previously overlooked in this poorly studied area characterized by an arid bioclimate.
In the locality of Bouarfa, we found that M. crassus was living in sympatry with another Gerbilline (Tarabul’s gerbil Gerbillus tarabuli Thomas, 1902). And in the locality of Ain Beni Mathar, we captured M. crassus just a few kilometres from a small colony of M. shawii inhabiting a cultivated and irrigated area with cereal crops. Generally, M. crassus is not regarded as sympatric with the other jird species in North Africa at the macrohabitat level (Petter, 19616; Aulagnier et al., 2017), but it seems to cohabitate with M. libycus and M. shawii in the Oriental region of Morocco, possibly through microhabitat segregations. This strategy may be beneficial to avoid interspecific competition (Shioya et al., 1990). Many studies have explained segregation patterns of coexisting rodent species in terms of multidimensional structure distinction of microhabitats. For example, the desert kangaroo rats, Dipodomys ordii Woodhouse, 1853 and Dipodomys merriami Mearns, 1890, overlap their habitats significantly but avoid direct competition by means of subtle differences in habitat preference (Schroder and Rosenzweig, 1975). In another example, habitat segregation allows the coexistence in sympatry of Jaculus jaculus Linnaeus, 1758 and Jaculus hirtipes Lichtenstein, 1823, two closely related species of desert jerboas living in North Africa (Shenbrot et al., 2016). Future ecological studies should address microhabitat occupation and interspecific competition in jirds and other Gerbillinae, particularly in the increasingly anthropized and agricultural landscapes of North Africa.
Finally, the Old World Cutaneous Leishmaniasis caused by Leishmania major Yakimoff and Schokhor, 1914, is a widespread parasitic disease that is transmitted by sand flies among a limited number of Gerbillinae reservoir-species. In Morocco, this disease occurs mainly in the south and east of the country and M. shawii is considered to be its main reservoir host (Rioux et al., 1982; Rhajaoui, 2011). However, the disease causative agent was found in M. crassus in the Sinai Peninsula (Morsy et al., 1991), Jordan (Amr, 2000) and Israel (Tsurim et al., 2020), although the Meriones individuals used in these studies could have been misidentified. Field surveys in search of the Gerbillinae host of the cutaneous leishmaniasis were carried out in the 1980s in the Tata region in southern Morocco, which was then home to an outbreak of the disease, but the captured M. crassus individuals were not infected (Rioux et al., 1982, 1984). Further work on M. crassus should examine its potential role in the transmission of this parasitic disease in Morocco and North Africa.
Acknowledgements
We thank Loubna Tifarouine (Mohammed V University in Rabat) for help in collecting the specimens. We are grateful to the three reviewers for their valuable comments and suggestions that have helped improve this paper.
















