
Arxius de Miscel·lània Zoològica. Volume 18 (2020) Pages: 101-111
Abundance and community structure of birds breeding in Kalahari woodland used as rangeland
Kopij, G.
DOI: https://doi.org/10.32800/amz.2020.18.0101Download
PDFCite
Kopij, G., 2020. Abundance and community structure of birds breeding in Kalahari woodland used as rangeland. Arxius de Miscel·lània Zoològica, 18: 101-111, DOI: https://doi.org/10.32800/amz.2020.18.0101-
Reception date:
- 02/06/2020
-
Acceptation date:
- 21/07/2020
-
Publication date:
- 10/09/2020
-
Share
-


-
Visits
- 4056
-
Downloads
- 1337
Abstract
Abundance and community structure of birds breeding in Kalahari woodland used as rangeland
The line transect method was applied to assess the linear index of abundance and community structure of birds breeding in an area of Kalahari woodland used as range land for cattle. Four transects were designed along roads traversing the ranch. Each transect was 2.5 km long and was surveyed three times in 2014 and 2015. A total of 47 bird species were recorded but only 25-31 species were recorded on any particular transect. In the neighbouring pristine Kalahari woodland, the number of bird species on a12 km long transect was much higher (n = 88), and ranged in some sections (each 1.2 km in length) from 35 to 53. On any particular section, the number of dominant species ranged from 5 to 7, and their cumulative dominance on each section was similar, ranging from 53-56 %, while the community dominance index ranged from 0.23 to 0.35. The most numerous species were the Cape turtle dove and emerald-spotted wood dove, which were dominant on all sections. Together they comprised 28 % of all breeding birds. The avian community in the Kalahari woodland in Sachinga LDC did not differ from the neighbouring pristine woodland either in terms of species diversity or evenness. However, it differed significantly in terms of species richness, the proportion of main ecological guilds, and linear index of abundance of particular species. These differences could be mainly due to the structure of woody vegetation, which is much thicker in Sachinga than in the pristine woodland not used as pasture for cattle. The thickening of this vegetation on the ranch could have been caused by heavy grazing pressure by the cattle.
Data published through GBIF (Doi: https://doi.org/10.15470/rova7r)
Key words: Community ecology, Population densities, Rangeland management
Resumen
Abundancia y estructura comunitaria de aves nidificantes en la sabana arbolada del Kalahari utilizada como pastizal
Se empleó el método del transecto lineal para evaluar el índice lineal de abundancia y la estructura comunitaria de aves nidificantes en la sabana arbolada del Kalahari utilizada como pastizal para ganado. Se definieron cuatro secciones de un transecto a lo largo de varios caminos que cruzan la finca. Cada sección tenía una longitud de 2,5 km y se estudió tres veces en 2014 y 2015. Se registraron un total de 47 especies de aves en el transecto, pero tan solo de 25 a 31 en una sección específica del mismo. En la zona inalterada colindante de sabana arbolada del Kalahari, el número de especies de aves registradas en el transecto de 12 km de longitud fue mucho más elevado (n = 88), variando desde 35 hasta 53 en secciones concretas (cada una de 1,2 km de longitud). En la sección específica, el número de especies dominantes osciló de 5 a 7 y su dominancia acumulada en cada sección fue similar, variando entre el 53 y el 56 %, mientras que el índice de dominancia comunitaria osciló de 0,23 a 0,35. Las especies más numerosas fueron la tórtola de El Cabo y la palomita aliverde, dominantes en todas las secciones y que en conjunto constituyen el 28 % de todas las aves nidificantes. La comunidad de aves de la sabana arbolada del Centro de Desarrollo de la Ganadería (LDC) de Sachinga no difiere de la existente en la zona inalterada colindante en cuanto a diversidad de especies ni homogeneidad. Sin embargo, difiere de forma significativa por lo que respecta a la riqueza de especies, la proporción de grupos ecológicos principales y el índice lineal de abundancia de especies concretas. Estas diferencias pueden deberse principalmente a la estructura de la vegetación forestal, que es mucho más densa en Sachinga que en la sabana arbolada colindante inalterada (no utilizada como pastizal para ganado). La frondosidad de esta vegetación puede haber sido causada por la elevada presión del pastoreo.
Datos publicados en GBIF (Doi: https://doi.org/10.15470/rova7r)
Palabras clave: Comunidad ecológica, Densidad de población, Gestión de pastizales
Resum
Abundància i estructura comunitària d’ocells nidificants a la sabana arbrada del Kalahari utilitzada com a pastura
Es va fer servir el mètode del transsecte lineal per avaluar l’índex lineal d’abundància i l’estructura comunitària d’ocells nidificants a la sabana arbrada del Kalahari utilitzada com a pastura per al bestiar. Es van definir quatre seccions d’un transsecte al llarg de diversos camins que travessen la finca. Cada secció tenia una longitud de 2,5 km i es va estudiar tres vegades els anys 2014 i 2015. Es van registrar un total de 47 espècies d’ocells en el transsecte, però només de 25 a 31 en una secció específica d’aquest. A la zona inalterada contigua de sabana arbrada del Kalahari, el nombre d’espècies d’ocells registrats en el transsecte de 12 km de longitud va ser molt més alt (n = 88), des de 35 fins a 53 en seccions concretes (cadascuna d’1,2 km de longitud). A la secció específica, el nombre d’espècies dominant va oscil·lar de 5 a 7 i la dominància acumulada en cada secció va ser similar, amb una variació del 53 al 56%, mentre que l’índex de dominància comunitària va oscil·lar de 0,23 a 0,35. Les espècies més nombroses van ser la tórtora del Cap i la tortoreta d’ales maragda, dominants en totes les seccions i que en conjunt constitueixen el 28 % de tots els ocells nidificants. La comunitat d’ocells de la sabana arbrada del Centre de Desenvolupament de la Ramaderia (LDC) de Sachinga no difereix de la que hi ha a la zona inalterada contigua pel que fa a diversitat d’espècies ni homogeneïtat. Tanmateix, difereix de manera significativa pel que fa a la riquesa d’espècies, la proporció de grups ecològics principals i l’índex lineal d’abundància d’espècies concretes. Aquestes diferències poden obeir principalment a l’estructura de la vegetació forestal, que és molt més densa a Sachinga que a la sabana arbrada contigua inalterada (no utilitzada com a pastura per al bestiar). La frondositat d’aquesta vegetació pot haver estat causada per l’elevada pressió de la ramaderia.
Dades publicades a GBIF (Doi: https://doi.org/10.15470/rova7r)
Paraules clau: Comunitat ecològica, Densitat de població, Gestió de pastures
Introduction
In all parts of Africa, the expansion and intensity of agriculture pose a major threat to biodiversity through the conversion of natural habitats to agrocenosis, fragmentation and degradation. However, in some places, where selected biota were carefully surveyed in agrocenosis, it has been shown that they may also well preserve certain communities, for example birds (Kopij, 1998, 2006, 2013, 2015, 2018; Ratclife and Crowe, 2001; Bolwig et al., 2006; Humle, 2007; Mulwa et al., 2012; Nadng’ang’a et al., 2013). This is especially evident when traditional agriculture is established within woodlands and wet savannas.
One of the largest broadleaved savanna type forests in Namibia is the Kalahari woodland. It occupies about 600,000 km2 in Kavango West, Kavango East, Zambezi, and eastern parts of Ohangwena and Oshikoto regions (Mendelsohn et al., 2009). This ecoregion supports a rich and diverse fauna that includes large populations of elephant Loxodonta africana, black rhino Diceros bicornis, white rhino Cerathotherium simum, leopard Panthera pardus, and wild dog Lycaon pictus (G. Kopij, own data). Nearly 500 bird species have been recorded in this biome (Kopij, 2016, 2017). Due to the scarcity of surface water, until rather recently, the Kalahari woodland was sparsely occupied by people, but in recent decades, human populations have been increasing and the cattle industry is growing, with far-reaching effects on the environment and wildlife (Mendelsohn et al., 2009). Every year, at least 1,000,000 m3 of raw timber harvested from this forest is shipped from Walvis Bay port to China (G. Kopij, own data). It is therefore essential to closely monitor and measure the resulting threats to the biodiversity and the whole ecosystem. This may help to formulate strategies and policies to protect this precious biome from widespread fragmentation, destruction, and degradation.
The purpose of this study was to estimate population densities and dominance of bird species occurring in Kalahari woodland used for cattle grazing and to compare these ecological parameters with the same habitat that remains in a pristine stage.
Material and methods
Study area
The study was conducted in the Sachinga Livestock Development Centre of the Ministry of Agriculture of the Namibian Government. The centre is situated in NE Namibia, in the Zambezi region, about 40 km west of Katima Mulilo (fig. 1), between 17º 39′ 40”-17º 40′ 20” S and 24º 23′ 55”-24º 24′ 02” E. The Centre is a ranch of 3,243 ha. The aim of the centre is in situ conservation, livestock improvement programmes and provision of breeding animals to the farmers, and farmer education. The centre is surrounded by communal areas with subsistence farming.
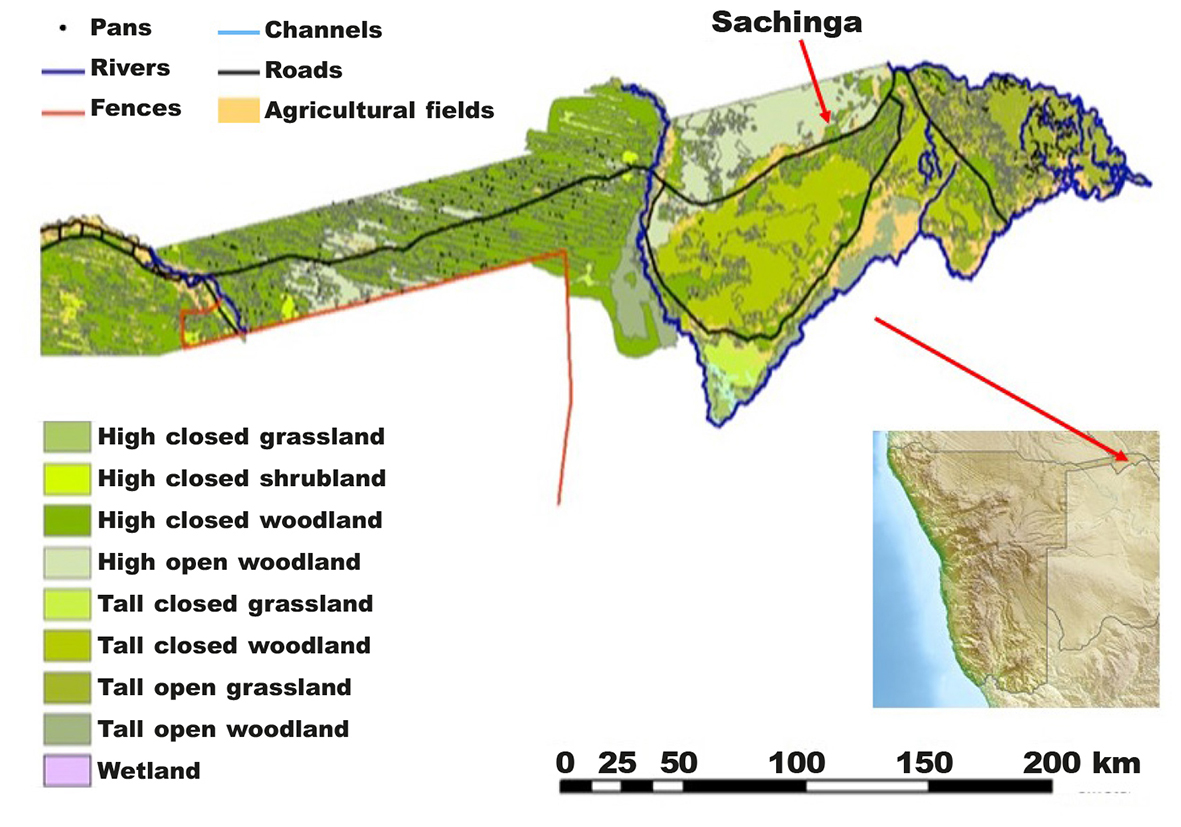
Fig. 1. Location of the study plot (Sachinga) within the Zambezi Region in NE Namibia. Fig. 1. Localización de la parcela de estudio (Sachinga) en la región de Zambeze, en el noreste de Namibia.
Most of the Sachinga LDC is covered by teak woodland, with smaller portion of Burkea-Kisat-False Mopane woodland and Omuramba grassland. The natural vegetation is well-preserved and represented by three classes of Kalahari woodland: 1) Colophospermum mopane, with two dominant associations: Nymphaea nouchali–Eragrostis rotifer which contains water species, and Combretum imberbe–Acacia nigrescens in association with sparse woodlands; 2) Baikiaea plurijuga woodland, comprising Guibourtia coleospherma–Burkea africana Association and Combretum collinum–Boscia albitrunca association; and 3) Baikiaea plurijuga–Terminalia sericea class with Pterocarpus angolensis–Diploryhnchus condylocarpon association, Dalbergia martinii–Berchemia discolor association, and Acacia fleckii–Baphia massaiensis association (Lushetile, 2009).
Methods
The American version of the transect line method (Bibby et al., 2012; Sutherland, 1996) was used to assess the population density and community structure of birds occurring in the ranch. A transect of 10 km in length and 100 m width (50 m on each side) was designed along roads traversing the ranch. The transect was divided into four equal sections (fig. 2). Each section was 2.5 km in length, and was surveyed three times (03/09/2014. 07/03/2015, 29/04/2015), each time in the same direction. Counts were conducted by the same observer, in the mornings, and in calm and cloudless weather conditions. Dominance is expressed as the percentage of the total number of pairs of a given species in relation to the total number of all pairs of all species recorded. A dominant species is defined as that comprising 5 % or more of all individuals of all species recorded, and a subdominant species is defined as that comprising 2-4.99 %.
Fig. 2. Transect section design (A, B, C, D) in the Sachinga Livestock Development Centre. Fig. 2. Diseño de las secciones de transectos (A, B, C, D) en el Centro de Desarrollo de la Ganadería de Sachinga.
All birds showing breeding (e.g. transporting nesting material, constructing nests, feeding chicks etc.) or territorial (e.g. singing males) behaviour were recorded. Special attention was paid to simultaneously singing males, as they were important in determining the number of occupied territories. Special care was taken not to count the same birds twice, as this could overestimate the number of territories.
The number of breeding pairs was estimated for each section on each transect. The maximumnumber of breeding pairs at each survey and on each section was assumed as the real number of breeding pairs. The total number of breeding pairs of each species on a particular transect was calculated as the total of maximum numbers recorded on each section within the transect.
The following guilds were distinguished: diet (G, granivorous; I, insectivorous; F, frugivorous; N, nectarivorous) and nesting (T, in trees or shrubs; H, in holes; G, on the ground; V, in herbaceous vegetation).
The following indices were used to characterise diversity, evenness and similarity between the communities:
1) Shannon’s diversity index: H’ = -∑ pi ln pi where: pi is the proportion of breeding pairs belonging to the ith species.
2) Simpson’s diversity index: D = ((∑ n (n – 1))/N (N – 1) where a :is the total number of breeding pairs belonging to a given species, N is the total number of breeding pairs of all species.
3) Pielou’s evenness index: J’ = (-∑ pi ln pi) / ln S where pi is the proportion of breeding pairs belonging to the ith species; S is the total number of species. J’ varies between 0 and 1. The lower the variation between species in a community, the higher the J’.
4) Community dominance index: DI = (n1 + n2) / N where n1, n2 are the number of pairs of the two most abundant species, N is the total number of pairs of all species.
5) Sörensen’s Coefficient: I = 2C / A + B where A is the number of bird species in one plot, B is the number of bird species in another plot, C is the number of bird species common to both plots.
Systematics and nomenclature of bird species follow Hockey et al. (2005). Scientific names of bird species are listed in appendix 1 and data paper published through GBIF (Doi: https://doi.org/10.15470/rova7r).

Appendix 1. Results of transect counts of birds on range land in the Kalahari woodland in 2014/2015: p, number of breeding pairs; p/km, linear index of abundance (pairs / 1 km); dom, dominance. (Dominant species are indicated in bold). Apéndice 1. Resultados de los recuentos de aves por transectos en la sabana arbolada del Kalahari en 2014/2015: p, número de parejas nidificantes; p/km, índice lineal de abundancia (pareja / 1 km); dom, dominancia. (Las especies dominanes están indicadas en negrita).
Results and discussion
A total of 47 bird species were recorded on Sachinga ranch (10 km), but only 25-31 were recorded on any particular section (2.5 km) of the transect (table 1, appendix 1, datapaper published through GBIF (Doi: https://doi.org/10.15470/rova7r). In the neighbouring pristine Kalahari woodland (Kopij, 2017), the number of bird species on the transect (12 km) was much higher (n = 88), and ranged in particular sections (1.2 km) from 35 to 53. The much lower number of species recorded in the ranch study area is probably mainly due to the fact that most of the vegetation on the transects was teak woodland and it showed signs of bush encroachment, which is avoided by many bird species. In Sachinga, March and April counts were conducted at the end of the rainy season, while in the pristine woodland, the October and November counts were conducted at the beginning of the rainy season (fig. 3). This difference could influence species richness, as birds are more common (in terms of the number of species) and more active at the onset of the rainy season than at the end of this period (own data).

Table 1. Characterisation of the breeding bird community in particular transect sections (A, B, C, D) in the Kalahari woodland managed as a ranch. Tabla 1. Caracterización de la comunidad de aves nidificantes en secciones de transectos concretos (A, B, C, D) en la sabana arbolada del Kalahari gestionada como rancho.

Fig. 3. Monthly rainfall in Katima Mulilo in 2014 and 2015. Fig. 3. Precipitación mensual en Katima Mulilo en 2014 y 2015.
The number of dominant species on each section ranged from 5 to 7, and their cumulative dominance on each section was similar, ranging from 53-56 %, while the Community Dominance Index ranged from 0.23 to 0.35. The most numerous species were the Cape turtle dove and emerald-spotted dove. Both species were dominants on all sections. Together they comprised 28 % of all breeding birds (appendix 1). Two other species, the fork-tailed drongo and the white-browed scrub robin, were dominant on 3 out of 4 sections. Four species, the crested Guineafowl, the orange-breasted bush-shrike, the grey-backed camaroptera and the swamp boubou, were recorded as dominant only on one section (appendix 1).
In the neighbouring Kalahari woodland in a pristine stage (Kopij, 2017), the mean number of species per section was much higher (n = 43), while the mean number of dominant species per section (3.8) and mean cumulative dominance (28.5 %) were much lower than in Sachinga. This difference is despite the fact that sections in the pristine woodland were shorter (1.2 km) than those in the ranch (2.5 km). The most numerous species in the pristine Kalahari Woodland was also the Cape turtle dove, and it was the only species which remained dominant on all 10 sections surveyed. Other species were recorded as dominant only on 1-3 of 10 sections surveyed. These groups included the emerald-spotted dove, the fork-tailed drongo, the swamp boubou, the yellow-bellied apalis, the white-browed scrub robin, the Cape starling, the black-chested prinia, and the long-billed crombec (Kopij, 2017).
The Simpson Diversity Index ranged from 0.95 to 0.97 on the section in the pristine Kalahari woodland (Kopij, 2017), and was much the same on the sections of transects in Sachinga, ranging from 0.93 to 0.95. The overall Evenness Similarity Index was also very similar in both habitats compared (0.88 in the pristine woodland vs. 0.84 in Sachinga woodland), with a similar range on the particular sections (0.86-0.97 vs. 0.88-0.91 respectively).
Although the overall linear index of abundance on the ranch and pristine Kalahari Woodland did not differ significantly (X2-test = 1,29, p > 0,05), the linear index of abundance of more numerous species (with at least 1 pair per 1 km of transect) differed markedly (table 2). Only the difference in the numbers of two species in this group, the fork-tailed drongo and the red-eyed dove, were not statistically significant.

Table 2. Comparison of the linear index of abundance (pairs /km) of the most numerous bird (> 1 pair / km) species on transects in the pristine woodland and the Kalahari ranch woodland. Tabla 2. Comparación del índice lineal de abundancia (parejas / km) de especies de aves más numerosoas (> 1 pareja / km) en transectos de la zona inalterada y en el rancho de la sabana arbolada del Kalahari.
The number of species that can be regarded as specialists, as rare, or of special conservation concern was much higher in the pristine woodland than in the ranch woodland. There were eleven raptor and owl species in the pristine woodland, but none were recorded in the ranch woodland. Six piciiform species were observed in the pristine woodland, compared to only three on the ranch, and 10 coraciiform species were observed n the pristine woodland compared to five in ranch woodland (table 2).
The differences between avian communities in the pristine and in the ranch woodlands were even more pronounced if September counts only are considered (tables 2 and 3). Although diversity indices were similar in these two Kalahari woodland areas, the numbers of species and numbers of breeding pairs were much higher in the pristine woodland.

Table 3. Characterisation of the breeding bird community in particular sections in the pristine Kalahari woodland (A, a) and in woodland managed as a ranch (B, b). In both habitats the total length of the sections was 7.5 km. Tabla 3. Caracterización de la comunidad de aves nidificantes en secciones concretas en la zona inalterada de la sabana arbolada del Kalahari (A, a) y en la zona gestionada como rancho (B, b). En ambos hábitats la longitud total de las secciones fue de 7,5 km.
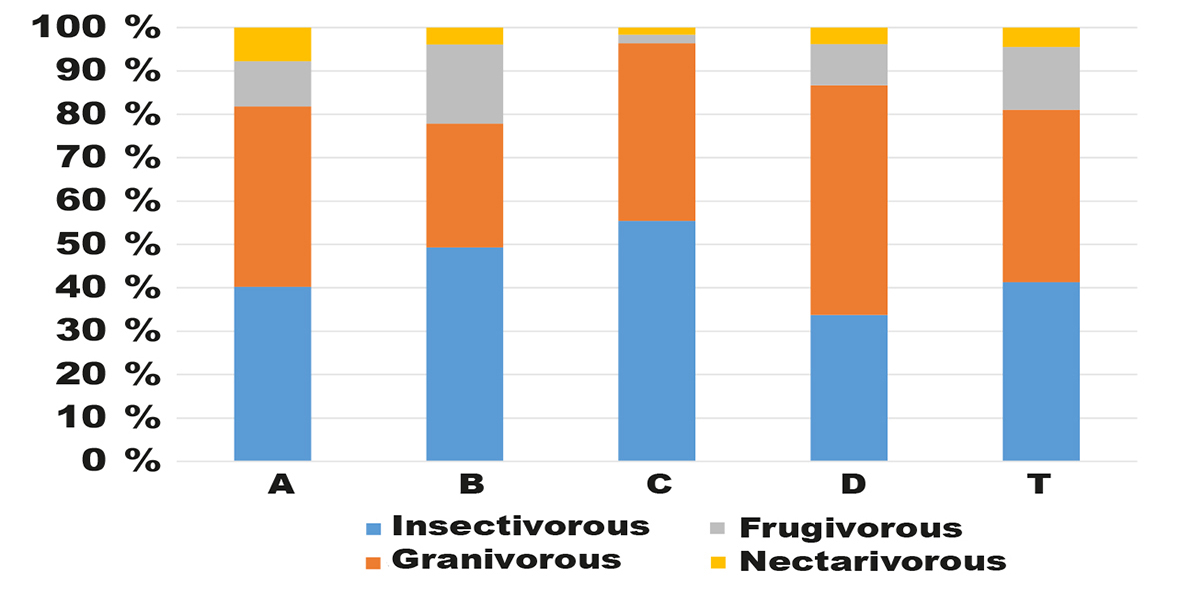
The contribution of insectivorous birds to the community in each section of the ranch woodland ranged from 33.8 % to 55.4 %, with a mean of 41.4 %. The proportion of granivorous birds was similar, ranging from 28.6 % to 52.9 %, with a mean of 39.9 % (fig. 4); in the pristine Kalahari woodland (Kopij, 2017), there were more insectivors (51.1 %), but fewer granivorous species (26.2 %) than on the ranch.
Fig. 4. Percent proportion of feeding guilds in each transect section (A, B, C, D) and in the whole transect (T). Fig 4. Porcentaje de grupos alimentarios en cada sección del transecto (A, B, C, D) y en todo el transecto (T).
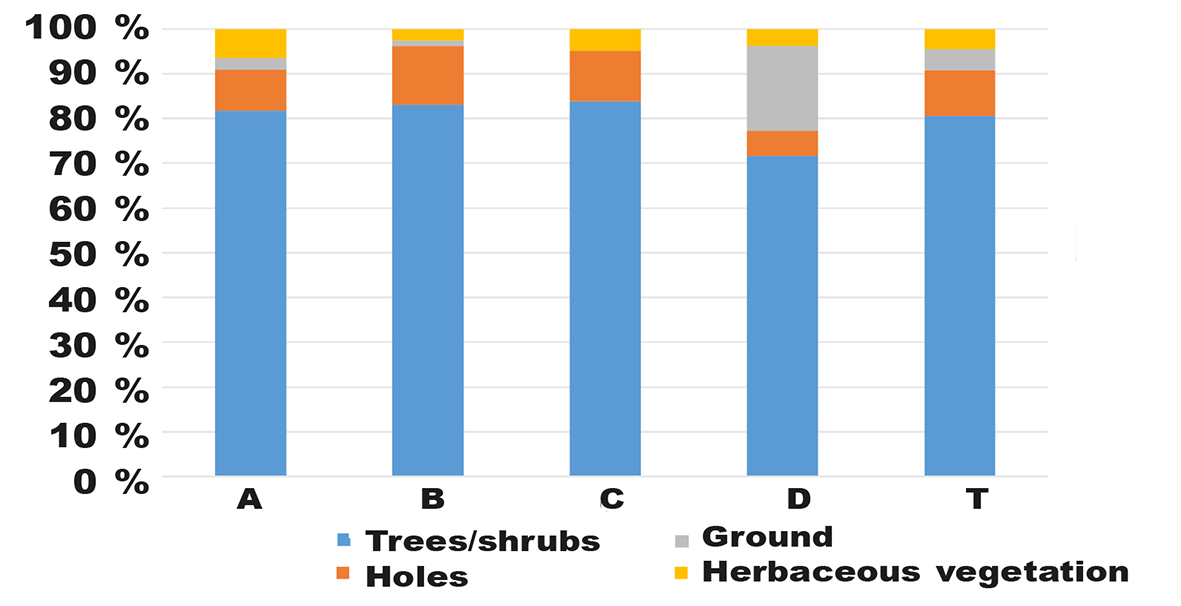
Nesting guilds in the ranch woodland were dominated (mean 80.6 %) by birds nesting in trees or shrubs. Hole-nesting birds comprised 10.1 % of all breeding birds (fig. 5). In the pristine woodland these guilds comprised 63.9 % and 28.0 %, respectively (Kopij, 2017).
Fig. 5. Percent proportion of nesting guilds in each transect section (A, B, C, D) and in the whole transect (T). Fig. 5. Porcentaje de grupos nidificantes en cada sección del transecto (A, B, C, D) y en todo el transecto (T).
In general, the avian community in the Kalahari woodland in Sachinga LDC does not differ from that in the neighbouring pristine woodland either in terms of species diversity or evenness. However, the community differs significantly in terms of species richness, the proportion of main ecological guilds, population densities of particular species, and the proportion of species of special conservation concern, both rare and specialist. These differences could be mainly due to the much thicker woody vegetation structure in Sachinga than in the neighbouring pristine woodland, which is not used as pasture for cattle. The thickening of this vegetation could have been caused by heavy cattle grazing.

















{kind=link}
{kind=link}
{kind=link}